
Role of adenosine in tumor progression: focus on A2B receptor as potential therapeutic target


Abstract
Adenosine receptors are a family of G-coupled receptors which mediate the anti-inflammatory and immune-suppressive effects of adenosine in a damaged tissue. A large number of evidence indicate that the accumulation of adenosine under hypoxic conditions favors tumor progression, helping cancer cells to evade immune responses. Tumor cells and/or lymphoid and myeloid cells can express the adenosine-generating enzyme CD73 and/or A2A receptor, which in turn strongly suppresses an effective T-cell-mediated response, while promotes the activity of suppressive cells such as Treg and myeloid-derived suppressor cells. CD73 inhibitors and A2A antagonists, either as single agents, or in combination with immune-checkpoints inhibitors such as anti PD-1 monoclonal antibodies, are currently in Phase I clinical trial in cancer patients. Recent studies show that A2B receptor plays an important role in mediating the pro-tumor effects of adenosine, since its selective blockade can inhibit tumor growth in some murine tumor models. Targeting A2B receptor reduces immunosuppression induced by myeloid cells and inhibits the stromal cells activity within the tumor microenvironment, limiting tumor angiogenesis and metastatic processes. Here, the authors review the current data on involvement of A2B receptor in regulating tumor progression and discuss the development of A2B receptor inhibitors as potential therapeutic agents in cancer treatment.
Keywords
Introduction
Tumor microenvironment is populated not only by malignant cells but also by other stromal cells and immune cells that cooperate to the development of cancer.[1,2]
In the eternal battle against cancer, several strategies have been developed. One of the first approach to treat cancer has been the antineoplastic chemotherapy which is made up of chemical substances that provide to halt directly the highly-replicating tumor cells by damaging their RNA or DNA.[3] Radiotherapy is another important treatment currently used for several tumors, through which cancer cells are either directly killed upon DNA damage by depositing high physical energy of radiation, or indirectly due to the release of free radicals.[4] Nowadays the most novel anti-cancer strategies are the targeted-therapy and immunotherapy. The cancer targeted-therapy uses small molecules that can block fundamental pathways or mutant proteins essential for tumor growth.[5] Conversely, cancer immunotherapy is a therapeutic strategy that improves the host immune response against cancer cells, instead of acting directly on tumor cells.[6]
Several chromosomal alterations, genetic mutations and genomic instability that occur in cancer cells provide a different set of antigens that the immune system can use to distinguish transformed cells from their own cells.[7] However, tumor cells escape from host immune surveillance through different mechanisms, that include loss of immunogenicity and ineffective T-cell mediated responses. Moreover, several inflammatory mediators including chemokines [CC-chemokine ligand 2 (CCL2), CCL5, CXC-chemokine ligand 1 (CXCL1), CXCL2, CXCL3, CXCL5, CXCL6, CXCL7, CXCL8, CXCL10 and CXCL12], cytokines [tumor necrosis factor (TNF), interleukin 1 (IL-1), IL-4, IL-5, IL-6, IL-10 and IL-13] and growth factors [granulocyte macrophage-colony stimulating factor (GM-CSF), vascular endothelial growth factor (VEGF), transforming growth factor-ß (TGF-ß)] are released by tumor cells and/or stroma and immune cells surrounding tumor tissue, generating a chronic inflammatory microenvironment. Chronic inflammation in cancer can facilitate tumor proliferation and invasion and drive the recruitment and activation of immunosuppressive cells, including T regulatory (Treg) cells, myeloid-derived suppressor cells (MDSCs) and tumor-associated macrophages (TAM). In this context, many inhibitory receptors, known as “immune checkpoint molecules” such as cytotoxic T-lymphocyte-associated antigen 4 (CTLA4) and programmed cell death-1 (PD-1), are upregulated on activated lymphocytes during an active immune response providing a negative feedback mechanism.[7] CTLA4 binds to members of the B7 family on antigen-presenting cells (APCs) inhibiting T-cell activation, while PD-1 interacts with ligands PD-L1, expressed on different cell types including tumor cells, or PD-L2 on macrophages and dendritic cells, inhibiting T-cell functions.[7,8] The development of agonist antibodies (for costimulatory pathways) or antagonist antibodies (for inhibitory pathways) which target lymphocyte receptors or their ligands is one of the most promising approach with the potential to modulate the tumor microenvironment and improve the efficacy of immune response/s against cancer cells.[8] The first class of immunotherapeutics approved by US Food and Drug Administration (FDA) for patients with metastatic melanoma includes antibodies against CTLA4 (Ipilimumab and Tremelimumab).[9] Later on other immune checkpoint molecules have been discovered, such as antibodies against PD-1 (Nivolumab, Pembrolizumab and Atezolizumab), PD-L1, lymphocyte-activation gene 3 (LAG3, also known as CD223), B7-H3 (also known as CD276), B7-H4 (also known as B7-S1, B7x and VCTN1) and T-cell immunoglobulin domain and mucin domain 3 (TIM3).[8] The therapeutic outcomes in cancer patients is improved by combining immunotherapeutics with chemotherapy.[8] The concomitant blockade of different immune checkpoints may increase the success of immunotherapy in cancer patients.[8] Hence, in the last few years many efforts have been made aiming to investigate novel therapeutic strategies to inhibit cancer-induced immune-suppression. It has become clear that in the tumor microenvironment there are several pathways that may play an important role in the tumor immune evasion process. Among them, extracellular adenosine, an ATP-derived molecule generated by the extracellular CD39/CD73 enzymes, has been identified as an immune checkpoint that critically impairs the anti-tumor immune response mainly via A2A adenosine receptor subtype.[10-12] Accordingly, selective inhibitors of adenosine signaling pathways have been tested in pre-clinical studies[13,14] and some of them, including the antibody anti-CD73 and the A2A receptor antagonists are currently in Phase I clinical trials in cancer patients, either as single agents, or in combination with immune checkpoints inhibitors such as anti PD-1 monoclonal antibodies [NCT02503774 and NCT02655822].
While the role in tumor immunity of CD73-A2A receptor axis has been extensively examined, less is known about the role of A2B receptor subtype in tumor development and progression. Compelling evidence suggest that this receptor contributes to the pro-tumor effects of adenosine within tumor microenvironment. In this article, we review the current data on the effects of adenosine in tumor progression, focusing on the emerging role of A2B receptor in regulating tumor growth and discuss the therapeutic potential of targeting A2B receptors in cancer treatment.
Critical roles of adenosine in tumor progression
Adenosine is a key endogenous molecule produced at the extracellular level by two ectoenzymes, ecto-5’-nucleotidase (CD73) and ectonucleoside triphosphate diphosphohydrolase-1 (CD39) physiologically expressed on both hematopoietic and non-hematopoietic cell types.[15,16] Once released in the extracellular space, adenosine elicits its physiological responses by coupling and activating four membrane adenosine receptors (A1, A2A, A2B, A3) which contain seven transmembrane domains coupled to G proteins.[17] Once bounded with its receptors, adenosine can inhibit (via A1 and A3) or stimulate (via A2A and A2B) the adenylyl cyclase resulting in a decrease or increase in cyclic AMP (cAMP) accumulation, respectively.[18] cAMP activates the protein kinase A (PKA) and in turn the nuclear substrate cAMP responsive element-binding protein (CREB) that regulates the expression of several genes by binding to cAMP responsive elements and other cAMP effectors such as Epac, altering pro-inflammatory genes expression.[19-22] A2B receptor can also activate the phospholipase C (PLC) by coupling Gq protein.[23,24] All adenosine receptors are involved in the modulation of mitogen-activated protein kinase (MAPK) activity.[25]
The tumor milieu is characterized by high levels of adenosine triphosphate (ATP) due to the high proliferating rate of cancer cells. The ATP is rapidly converted at the extracellular level in ADP and AMP through two reversible steps via CD39, while the last irreversible step in adenosine is mediated by CD73.[16] Under homeostatic conditions adenosine level is low but during pathophysiological events (including stress, infection, inflammation and cancer) extracellular adenosine levels can be increased from 10-200 nmol/L up to 10-100 μmol/L.[26] In inflammatory-associated conditions, adenosine typically attenuates the inflammatory response.[26,27] Importantly, studies by Ohta and Sitkovsky[28] showed for the first time that A2A receptor-deficient mice are unable to control inflammation, resulting in exaggerated immune responses which can trigger extensive tissue disruption with subsequent cell death. These effects of adenosine are dependent on the activation of the adenosine A2A receptors on immune cells, which induce a wide range of singular immunosuppressive responses which regulate the uncontrolled inflammation to harmful insults.[27,29,30] However, in the context of tumor while extracellular ATP increases the T-cell mediated effector function, high levels of adenosine mediates opposite effects favoring immune suppression that is associated with tumor growth and metastasis.[31] Hypoxia, which is a common feature of the tumor microenvironment that promotes immunosuppression, is one of the main factors responsible of the increased production of adenosine within many solid tumors.[11] Indeed, the expression and the enzymatic activities of CD39 and CD73, responsible of the adenosine generation, increased under hypoxic conditions, while the expression of the adenosine kinase, which inhibits the metabolism of adenosine, is down-regulated.[11] At the same time, the expression of adenosine receptors A2A and A2B is also up-regulated.[32] Consequently, adenosine along with other HIF-induced immunosuppressive factors and cells, contributes to modulate the functions of tumor cells, tumor-infiltrating immune cells and/or other stroma cells.
Within the hematopoietic compartment, CD39 is expressed on B cells and monocytes, subsets of CD8+ T cells, CD4+ T cells and NK cells.[15,33] CD73 is expressed on B cells and subsets of CD8+ T cells, CD4+ T cells and NK cells and small subsets of monocytes.[15,33] CD39 and CD73 are co-expressed on B cells, Treg cells, Th17 cells, NK cells, neutrophils, tissue macrophages and myeloid-derived suppressor cells (MDSCs).[15,33] CD39 and CD73 are also expressed on endothelial cells and on the surface of several types of cancer cells.[14,15] Thus, CD73-expressing cells, including immune cells and/or stroma cells, produce adenosine that accumulate in the tumor microenvironment and profoundly impairs anti-tumor immune responses. Accordingly, a large number of evidence have proved that targeting adenosine-generating enzymes significantly reduces tumor growth by improving anti-tumor immune responses.
A2A receptor is the most thoroughly characterized receptor involved in the adenosine-induced anti-inflammatory/immune-suppressive effects within the tumor microenvironment. A2A receptor is highly expressed on lymphocytes, macrophages, dendritic cells, NK cells, and neutrophils. Activation of A2A receptor significantly reduces T-cell receptor (TCR)-triggered effector functions, including proliferation and production of cytokines and chemokines, preventing T cells activation and function via cAMP/protein kinase cAMP-dependent (PKA) pathways.[34-36] These effects occurs upon A2A adenosine receptor stimulation in naïve CD4+ T cells as well as in CD8+ T cells. Furthermore, A2A receptor stimulation reduces the expression of CD25 and CD40 ligand (CD40L) and increases the expression of PD-1 and CTLA-4 on T cells,[37] inducing T cell anergy that promotes peripheral tolerance.[35] Stimulation of A2A receptor on myeloid cells can also affect the release of IL-12 and induce the production of IL-10,[38] affecting significantly the T- and NK-cell responses in the solid tumor microenvironment.[39] Additional evidence also show that A2A adenosine receptor stimulation promotes the development of immune suppressive myeloid cells[40] or Treg cells.[41] The first in vivo genetic evidence of the role of A2A receptor in tumor progression has been reported by Ohta et al.[10] who showed that 60% of A2A receptor deficient mice completely rejected established immunogenic tumors in a CD8+ T-cell-dependent manner. However, 40% of tumor-bearing A2A receptor deficient mice did not reject the tumor, possibly because of the expression of A2B receptor on A2A receptor deficient CD8+ T cells.[10]
At the same time a large number of evidence show that inhibition of CD73 activity or CD73 knockdown on tumor cells inhibit tumor growth and metastasis by enhancing the anti-tumor T cell response.[42-46] CD73-deficient mice are resistant to tumor and show an increased influx of CD8+ T cells[44] and low number of Tregs within the tumor.[47]
The expression of CD73 on various tumor cells from cancer patients, including breast,[46] glioblastoma,[48] prostate,[49] ovarian,[50] leukemia[51] has been associated with poor prognosis. Notably, some chemotherapeutics are able to increase the expression of CD73 on cancer cells, which may in turn represent a putative mechanism of resistance to chemotherapeutics.[46,49,51] On the other hand, targeting CD73 can improve the therapeutic potential of some conventional cancer treatments, including chemotherapy, radiotherapy and immunotherapy. For example, inhibition of CD73 in combination with doxorubicin prolonged the survival of mice with metastatic breast cancer.[46] Adenosine can also impair the anti-tumor response induced by high dose of radiation therapy.[52] Administration of CD73 inhibitor into mice with tumors exposed to radiation therapy can significantly reduce tumor growth.[52] Notably, inhibition of CD73 may also improve the synergy of radiation therapy in combination with anti-CTLA4 monoclonal antibody.[52]
Recent studies indicate that inhibition of adenosine/A2 adenosine receptors axis synergizes with other immune checkpoints inhibitors reducing potently tumor growth in murine models of cancer. In particular, treatment of mice with monoclonal antibody anti-CD73 enhances the anti-tumor effects of antibodies anti-PD1 and anti-CTLA4.[53] In support, other studies have demonstrated that selective blockade of A2A adenosine receptor in combination with anti PD-1 antibody and anti-CTLA4 antibody potently reduced tumor growth.[54-56] The therapeutic synergy of these combinations depends on the CD73 expression on tumor cells, proving that CD73-generating adenosine by tumor cells within the tumor microenvironment may affect the activity of immunotherapy. Furthermore, blockade of PD-1 enhances the expression of A2A receptors on tumor-infiltrating CD8+ T cells, suggesting that adenosine via A2A receptor limits the immune response against cancer induced by inhibitors of immune checkpoints.[54] More recently, it has been demonstrated that blockade of A2B adenosine receptor subtype with a selective antagonist improves survival and the anti-metastatic effects of anti-PD1 and anti-CTLA4 monoclonal antibodies in both melanoma and mammary cancer models of metastasis with cells expressing CD73.[57] The anti-metastatic effects of these combinations relies on the capacity of immune checkpoints inhibitors to boost immune responses and on direct effects of A2B adenosine receptor inhibitor on cancer cell metastasis.[57] Here the authors show that blockade of A2B receptor in A2B receptor deficient mice is able to reduce the metastasis of human triple negative breast cancer (TNBC) xenografts, confirming the critical role of A2B receptor on cancer cells rather than host cells.[57] Altogether these preclinical studies strongly support the therapeutic potential of targeting adenosine in cancer.
Experimental evidence suggests that also CD39 can represent a potential therapeutic target for cancer treatment. CD39 is highly expressed by Treg cells and together with CD73 generate adenosine in the tumor microenvironment.[58] Elevated levels of CD39-expressing Treg cells have been found in some mouse tumor tissues, including melanoma and colorectal cancer.[58] Inhibition of CD39 reduces the tumor growth, enhances the recruitment of T cells in the tumor lesions and improves the effector functions of CD8+ T cells and NK cells, by impairing the activity of CD39-expressing Treg cells.[58] Although additional studies are needed to better clarify the therapeutic potential of targeting CD39 in cancer, the use of CD39 inhibitors might be useful to limit the immune suppression induced by Treg cells.
Selective agonists of A3 adenosine receptor subtype have proved to directly inhibit proliferation of A3-expressing tumor cells by arresting cell cycle progression and exert immunostimulatory effects in some murine tumor models in a NK- and T-cell-dependent manner, enhancing the production of Th1-like cytokines in the tumor microenvironment.[59-63] A3 adenosine receptor agonists have been tested indeed in some clinical trials for rheumatoid arthritis (NCT00280917, NCT00556894, NCT01034306, NCT02647762),[64] hepatocellular carcinoma (NCTNCT00790218, NCT02128958) and hepatitis (NCT00790673),[65] dry eye syndrome (NCT01235234, NCT00349466)[66] and psoriasis (NCT01265667).[67]
Nonetheless, emerging evidence suggest that A2B receptor can mediate the pro-tumor effects of adenosine. It is known that A2B receptor is important in some patho-physiological conditions, including vascular injury,[68] chronic lung disease,[69] vascular leak,[70] and ischemic disease.[71] First studies performed by Ryzhov et al.[72] in 2008 show that tumor growth in A2B receptor deficient mice was reduced compared to that observed in wild type mice, providing the first genetic evidence for a pivotal role of A2B receptor in tumor progression.
Up to now a number of selective A2B receptor antagonists (such as MRS1754, ATL801, GS-6201, PSB603 and PSB1115) and selective A2B agonists (Bay60-6583) have been synthesized, helping the study and the characterization of the role of this adenosine receptor in many patho-physiological conditions, including cancer, as discussed below.
Expression of A2B receptor
A2B adenosine receptor is widely expressed in the entire organism, although its role is not completely understood. The A2B receptor expression has been detected in type II alveolar epithelial cells,[73] endothelial cells,[74] chromaffin cells,[75] astrocytes,[76] neurons,[77] and taste cells.[78] Moreover, A2B receptor is expressed also on many immune cell populations including mast cells,[79] neutrophils,[70] dendritic cells,[80] macrophages,[74] and lymphocytes.[81]
Despite A2B receptor binds adenosine with lower affinity (EC50 = 24 μmol/L) than A2A receptor,[72,82] its relevance in regulating tumor growth is becoming clear both because its expression is highly influenced by the tumor milieu and because A2B receptor can play different physiological roles compared to A2A receptor.
The tumor microenvironment is characterized by high proliferating rate of cancer cells which contribute to hypoxia condition. Hypoxia is a very strong stimulus for up-regulating A2B receptor expression through hypoxia inducible factor (HIF-1α) and hypoxia-dependent signaling pathways in endothelial cells, dendritic cells (DCs), muscles, fibroblasts and T cells.[32,83-87] Indeed, a functional hypoxia-responsive region within the A2B receptor promoter has been identified, confirming the selective transcriptional induction A2B receptor by hypoxia.[87] Transcription of A2B receptor can be induced by bacterial lipopolysaccharide (LPS) or interferon (IFN)-γ in macrophages,[88,89] by TNF-α in vascular smooth muscle cells,[90] and by IL-1β in endothelial cells.[91] Furthermore, a post-transcriptional regulation of A2B receptor by inflammatory mediators has been demonstrated in endothelial and pulmonary epithelial cells[92] and in colonic epithelial cells.[93] Therefore, although A2B is a low-affinity adenosine receptor, under inflammatory-hypoxic conditions, its expression is up-regulated while the concentration of adenosine reaches highest levels. In this context, the A2B receptor may play an important role in mediating adenosine-induced pathological effects.
A2B receptor and tumor immunity
Although the role of A2B receptor in controlling T-cell-mediated response is not completely clear, compelling evidence indicate that this receptor may influence the features of some immune cell populations.
It has been demonstrated that A2B receptor is involved in the differentiation of T cells under Treg skewing-conditions, since its inhibition is able to suppress the expression of FoxP3 and IL-10 production in a way completely independent from T cell activation.[94]
To be activated and provide anti-tumor responses CD4+ T-cells need the expression of the major histocompatibility complex (MHC) class II. In several types of tumors, the loss of MHC class II is related to impaired levels of CD4+ T-cells.[95] Moreover, the levels of either MHC class II or class II transactivator (CIITA) are altered in highly metastatic cancer cells.[96] A2B receptor stimulation by repressing CIITA can impair MHC class II transcription in IFN-γ-stimulated cells.[97,98] Moreover, bone marrow-derived dendritic cells (BMDCs) express A2B receptor and adenosine inhibits BMDCs IL-12p70 production via A2B receptor. Depending on the levels of this cytokine, CD4+ T-cells can differentiate into Th1 or Th2 cells.[99] The impaired production of pro-inflammatory cytokines (TNF-α and IL-12) and the increased IL-10 production induced by A2B receptor activation leads to a lower expression of CD86 and MHC class II lowering CD4+ T cell stimulation.[100]
A2B receptor can also affect macrophages proliferation induced by macrophage colony-stimulating factor (M-CSF)[101] and the differentiation of human monocytes, mouse peritoneal macrophages and hematopoietic progenitor cells (HPCs) into myeloid DCs with tolerogenic and angiogenic features.[80] A2B receptor activation promotes the expansion in vitro of MDSCs, that contribute to induce immunosuppression by producing adenosine.[102] MDSCs potently suppress anti-tumor T-cell response and/or promote angiogenesis.[103] Altogether, these studies strongly support a role of A2B receptor in inducing the differentiation of hematopoietic progenitor cells into mature cells with tolerogenic and suppressive features. Subsequent studies performed in vivo show that A2B deficient mice have reduced amounts of tumor-infiltrating myeloid cells CD11bhigh/Gr-1high, suggesting that A2B receptor suppresses immune surveillance.[72] Later, Cekic et al.[104] showed that the selective blockade of A2B receptor inhibits bladder and breast tumor growth in mice, by inducing a T-cell mediated response in a CXCR3-dependent manner. In a mouse model of melanoma, selective blockade of A2B receptor inhibits tumor growth.[105] This effect was associated with lower levels of IL-10 and MCP-1 in the tumor tissue and reduced accumulation of tumor-infiltrating MDSCs.[105] Notably, the levels of MDSCs in secondary lymphoid organs remained unchanged in mice treated with the selective A2B receptor antagonist, consistent with a selective activity of the antagonist on the recruitment of MDSCs to tumor lesions rather than with a putative systemic effects.[105] Blockade of A2B receptor within the tumor microenvironment modulates the intra-tumoral levels of various inflammatory mediators and growth factors that could in turn influence the features of tumor-infiltrating immune cells, promoting the recruitment/accumulation of MDSC.[106] Accordingly, the percentage of tumor-infiltrating CD8+ T cells upon A2B receptor blockade enhanced in the tumor lesions.[105] Furthermore, treatment of mice with the A2B receptor antagonist PSB1115 in combination with dacarbazine, a chemotherapeutic agent commonly employed in melanoma patients, reduces tumor growth and significantly increases the number of CD8+ T cells in the melanoma lesions demonstrating the high potential of combining A2B receptor blockade and chemotherapy for cancer treatment.[105,106]
In conclusion, the experimental evidence in some tumor mouse models suggest that the selective blockade of A2B receptor may ameliorate T cell-mediated immune surveillance by impairing the accumulation of suppressive cells and the levels of inflammatory factors in the tumor microenvironment.[72,104-106] However, despite the relevance of these observations, more studies are needed to provide a detailed understanding of the role of A2B receptor in modulating the immune responses in tumor environments.
A2B receptor and tumor stroma
A number of studies indicate that A2B receptor can directly affect the proliferation/migration of tumor cells and the function of other stroma cells that populate the tumor niche, including endothelial cells and fibroblasts.
A critical role for A2B adenosine receptor in mediating proliferation and/or apoptosis in different cancer cell lines has been delineated. A2B adenosine receptor is highly expressed in prostate cancer cell lines and selective antagonist of A2B adenosine receptors or silencing A2B receptors blocked the proliferative effects induced by a non-selective adenosine analog NECA.[107,108] Other studies indicate that A2B adenosine receptor is highly expressed also in oral squamous carcinoma cell lines, as well as in human oral carcinoma tissues, where its expression is correlated with those of HIF-1.[109] Studies by Gessi et al.[110] demonstrate that in colon cancer cells, although at the mRNA levels A2B receptor is more expressed than A1, A2A and A3, the density of A3 receptors is the highest among the adenosine receptor subtypes. Later, other studies have demonstrated that the adenosine A2B receptor is up-regulated in colorectal carcinoma tissues and colon cancer cell lines compared with normal colorectal mucosa under hypoxic conditions.[111] Antagonists of A2B receptors inhibit cancer cell proliferation, suggesting that this receptor may be a potential therapeutic target for colorectal cancer.[111]
In contrast, in gastric cancer cells A2B adenosine receptor has been identified as target of miR-128b, a proto-oncogene miRNA down-regulated in gastric cancer tissues.[112] In this work, the authors demonstrate that the down-regulation of miR-128b in gastric cancer cell is associated with an over-expression of A2B adenosine receptor and decreased cell apoptosis rate.[112] In osteosarcoma cells it has been demonstrated that p73 upregulates A2B adenosine receptor and A2B receptor agonists can enhance p73-dependent cell death in response to chemotherapy.[113] Moreover, stimulation of A2B receptor with a non-selective adenosine analog NECA induces apoptosis in ovarian cancer cells.[114] Nonetheless,while a number of studies demonstrate that stimulation of A2B adenosine receptor in some cancer cell types promotes proliferation, whereby knockdown or pharmacological inhibition of this receptor reduces tumor cell growth and promotes apoptosis,[107-111] opposite results have been also described.[112,113] The discrepancy might likely depend on the cancer cell types, the expression levels of this receptor on tumor cells and the selectivity and/or concentrations of pharmacological tools used in the experimental settings.
It has been demonstrated that agonists of A2B receptor induce anti-proliferative and pro-apoptotic effects on glioblastoma cancer stem cells (CSCs).[115] Furthermore, stimulation of A2B receptors as well as A1 receptors sensitize glioblastoma CSCs to chemotherapy.[115]
A role of A2B receptor in promoting the migration of tumor cells in vitro and in vivo has been clearly demonstrated. Indeed, a number of studies show that adenosine may directly influence the migration/invasion of tumor cells via A2B adenosine receptor. Stagg et al.[42] have demonstrated that targeting the adenosine-generating enzyme CD73 inhibits tumor growth in mice and significantly delays the development of spontaneous lung metastasis.While the effect of anti-CD73 monoclonal antibody therapy on primary tumor growth relays on its capacity to improve immune surveillance, the anti-metastatic effects to the lungs is rather dependent on a direct effect of CD73-generating adenosine on breast tumor-cell migration via A2B adenosine receptors stimulation.[42] Consistent with the role of A2B receptor in promoting metastasis of breast cancer cells to the lung, administration of selective or non-selective A2B receptor antagonists into mice significantly reduced metastasis burden.[42,104] Furthermore, antagonists of A2B receptor preferentially inhibits the invasive capacity of breast cancer cells expressing Fos-related antigen-1 (Fra-1), a transcription factor overexpressed in human metastatic breast cancers.[116] Therefore, the authors suggest that Fra-1 activity is a prognostic indicator of both breast cancer metastasis and responsiveness to pharmacological inhibitors, such as A2B receptor antagonists.[116]
In a recent paper it has been demonstrated that high expression of A2B receptor is associated with poor survival in triple negative breast cancer (TNBC) patients.[57] As mentioned above, these authors demonstrate that A2B receptor antagonist prevents metastasis of A2B receptor-expressing tumor cells and improves survival when administered in combination with chemotherapeutic agents and immune checkpoints inhibitors monoclonal antibodies in both experimental and spontaneous murine models of metastasis.[57] The anti-metastatic effects of A2B receptor antagonists is independent on lymphocytes and myeloid cells, whilst tumor A2B receptor is critical.[57] These evidence highlight that A2B receptor may be an attractive target for treatment of breast metastasis.
A2B adenosine receptor can also contribute to the pro-angiogenic effects of adenosine in the tumor milieu. Vascular endothelial growth factor (VEGF) is a well-known mediator critically involved in tumor progression and angiogenesis.[117] A number of studies linked VEGF production to adenosine A2B receptor in human endothelial cells,[118,119] in some tumor cell lines[120,121] and in host immune cells, including dendritic cells and myeloid-derived suppressor cells.[72,80,122]
A2B receptor is expressed on human endothelial cells and its stimulation promotes the expression of several pro-angiogenic factors, including VEGF, IL-8 and basic fibroblast growth factor (bFGF).[118] Importantly, under hypoxic conditions the expression of A2B receptor in endothelial and smooth muscle cells increased and the stimulation of these receptors further enhance VEGF release.[119] Hypoxia is a common feature of tumor and can induce angiogenesis. At the same time, adenosine, whose levels became elevated during hypoxia, further enhances angiogenesis by stimulating A2B receptors, creating a positive feedback between hypoxia, adenosine and VEGF.
Other studies also indicate that adenosine promotes the release of angiogenic factors, namely VEGF and IL-8, in some cancer cells lines, via A2B receptor including human melanoma cells[120] and glioblastoma cells, which express high levels of A2B receptor under hypoxic conditions.[121]
Using A2B receptor deficient mice, Ryzhov et al.[72] firstly demonstrated the critical role of A2B receptor in modulating the VEGF levels in tumor tissues. Importantly, vascularization and tumor tissue VEGF levels were significantly reduced in A2B receptor deficient mice compared with WT mice.[72] This effect was associated with reduced tumor infiltration of VEGF-producing myeloid cells, suggesting that A2B receptor can modulate the release of VEGF either from tumor cells and from host tumor-infiltrating immune cells,[72] that can contribute to promote tumor angiogenesis.[123] As mentioned above, adenosine promotes the differentiation of dendritic cells precursors into a subset of DC that produce angiogenic factors, including VEGF, and other immunosuppressive factors via A2B adenosine receptor.[80] Notably, A2B-stimulated dendritic cells are able to promote tumor growth when injected into mice.[80] These observations strongly suggest that adenosine sustains tumor angiogenesis during tumor growth by stimulating the release of VEGF from endothelial cells, tumor cells and immune cells. Accordingly, targeting CD73 in mice impairs tumor angiogenesis and decreases VEGF levels, at least in part by lowering adenosine generation in tumor environment that activates A2B receptors.[124] Therefore, targeting CD73 and/or A2B receptor may represent a potential therapeutic strategy to block angiogenesis. In support of this, the pharmacological blockade of A2B receptor with a selective antagonist in mice significantly reduces the tumor levels of VEGF and CD31 positive cells within tumor lesions.[122] Moreover, the anti-angiogenic effect of A2B receptor antagonists is, at least in part, dependent on the lower frequency of tumor-infiltrating suppressive myeloid cells (MDSCs),[72,122] breaking the positive feedback loop that promotes angiogenesis and MDSC-mediated immune suppression in the tumor environment. Recent evidence indicate that A2B receptor stimulation promotes the release of FGF-2 and C-X-C motif chemokine ligand 12 (CXCL12) from tumor-associated fibroblasts,[125] that contribute to promote tumor growth and angiogenesis.[126] These effects are associated with reduced expression of fibroblast activation protein (FAP), a common marker of tumor-activated fibroblasts termed cancer-associated fibroblasts (CAF), that promote tumor growth enhancing tumor immune evasion and tumor vascularization.[127] A2B receptor-induced CXCL12 by tumor-associated fibroblasts contributes to the pro-angiogenic effects of A2B receptor via CXCR4, suggesting a link between tumor fibroblasts and endothelial cells.[127] Moreover, fibroblasts express CD73, which is up-regulated under hypoxic conditions.[127] Altogether, these evidence suggest that in the context of tumor A2B receptor contributes to mediate multiple effects of adenosine on different types of cells that populate the tumor niche. Furthermore, blockade of A2B receptor modulates the intra-tumoral levels of paracrine factors, which are critical in regulating intercellular crosstalk in the tumor microenvironment.
Although the predominant role of A2A receptor in mediating the immunosuppressive effects of adenosine in the tumor tissue and the high therapeutic potential of blocking adenosine generation and the A2A-mediated effects, by using anti-CD73 monoclonal antibodies and A2A selective antagonists, respectively, it is becoming clear that A2B receptor may significantly affect tumor progression and metastasis. Its contribute to tumor development and growth is most likely dependent on its high expression levels on tumor cells, and/or endothelial cells and/or other tumor-infiltrating cells, in a rich adenosine environment.
Conclusion
Adenosine plays a critical role in tumor immunity, angiogenesis and metastasis process. Strategies aimed to inhibit tumor adenosine production and functions, by using CD73 inhibitors and selective blockade of A2A adenosine receptor, are effective for cancer treatments, especially in combination with chemotherapeutic agents and immune-checkpoints inhibitors.
Nonetheless, compelling evidence support the role of A2B receptor subtype in contributing to the pro-tumor effects of adenosine within the tumor microenvironment, including immune suppression, angiogenesis and metastasis [Figure 1]. Despite these evidence, further studies are needed to better investigate thoroughly the mechanisms by which blockers of this receptor limit tumor growth. Understanding the relative role of A2B receptor in tumor, depending on the cell types, on its distribution and expression, will help to potentially apply A2B receptor-targeting agents for cancer treatment.
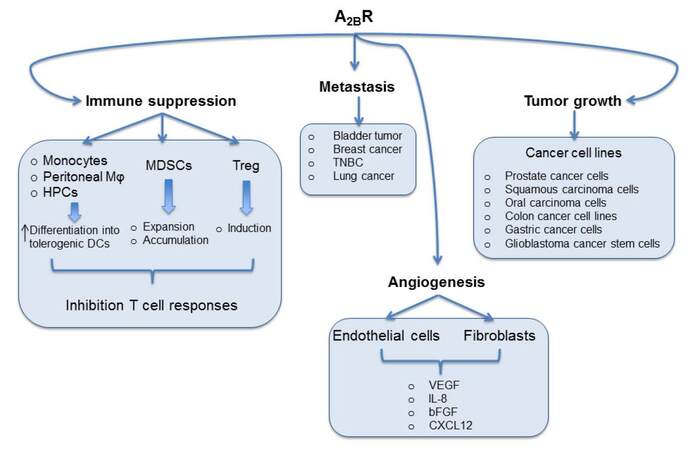
Figure 1. Multiple roles of A2B adenosine receptors in cancer. A2B receptor stimulation induces (1) the differentiation of human monocytes, mouse peritoneal macrophages (φ) and hematopoietic progenitor cells (HPCs) into tolerogenic dendritic cells (DCs); (2) the expansion and accumulation of MDSCs; (3) Treg differentiation, enhancing immune suppression that inhibits T-cell responses. Activation of A2B receptors on stroma cells, including tumor cells, endothelial cells and fibroblasts promotes tumor proliferation or invasion and angiogenesis. TNBC: triple negative breast cancer; VEGF: vascular endothelial growth factor; IL-8: interleukin-8; bFGF: basic fibroblast growth factor; CXCL12: C-X-C motif chemokine ligand 12; MDSCs: myeloid-derived suppressor cells
Authors’ contributions
Conceived and designed the study: C. Sorrentino, S. Morello
Performed literature search and prepared manuscript: C. Sorrentino, S. Morello
Revised the manuscript: S. Morello
Financial support and sponsorship
None.
Conflicts of interest
There are no conflicts of interest.
Patient consent
There is no patient involved.
Ethics approval
This article does not contain any studies with human participants or animals.
REFERENCES
1. Pietras K, Ostman A. Hallmarks of cancer: interactions with the tumor stroma. Exp Cell Res 2010;316:1324-31.
2. Klemm F, Joyce JA. Microenvironmental regulation of therapeutic response in cancer. Trends Cell Biol 2015;25:198-213.
4. Baskar R, Dai J, Wenlong N, Yeo R, Yeoh KW. Biological response of cancer cells to radiation treatment. Front Mol Biosci 2014;17:1-24.
5. Druker BJ, David A. Karnofsky Award lecture. Imatinib as a paradigm of targeted therapies. J Clin Oncol 2003;21:239-45s.
8. Pardoll DM. The blockade of immune checkpoints in cancer immunotherapy. Nat Rev Cancer 2012;12:252-64.
9. Hodi FS, O'Day SJ, McDermott DF, Weber RW, Sosman JA, Haanen JB, Gonzalez R, Robert C, Schadendorf D, Hassel JC, Akerley W, van den Eertwegh AJ, Lutzky J, Lorigan P, Vaubel JM, Linette GP, Hogg D, Ottensmeier CH, Lebbé C, Peschel C, Quirt I, Clark JI, Wolchok JD, Weber JS, Tian J, Yellin MJ, Nichol GM, Hoos A, Urba WJ. Improved survival with ipilimumab in patients with metastatic melanoma. N Engl J Med 2010;363:711-23.
10. Ohta A, Gorelik E, Prasad SJ, Ronchese F, Lukashev D, Wong MK, Huang X, Caldwell S, Liu K, Smith P, Chen JF, Jackson EK, Apasov S, Abrams S, Sitkovsky M. A2A adenosine receptor protects tumors from antitumor T cells. Proc Natl Acad Sci U S A 2006;103:13132-7.
11. Sitkovsky MV, Hatfield S, Abbott R, Belikoff B, Lukashev D, Ohta A. Hostile, hypoxia-A2-adenosinergic tumor biology as the next barrier to overcome for tumor immunologists. Cancer Immunol Res 2014;2:598-605.
12. Vaupel P, Mayer A. Hypoxia-driven adenosine accumulation: a crucial microenvironmental factor promoting tumor progression. Adv Exp Med Biol 2016;876:177-83.
13. Leone RD, Lo YC, Powell JD. A2aR antagonists: next generation checkpoint blockade for cancer immunotherapy. Comput Struct Biotechnol J 2015;13:265-72.
14. Allard B, Longhi MS, Robson SC, Stagg J. The ectonucleotidases CD39 and CD73: novel checkpoint inhibitor targets. Immunol Rev 2017;276:121-44.
15. Antonioli L, Pacher P, Vizi ES, Haskó G. CD39 and CD73 in immunity and inflammation. Trends Mol Med 2013;19:355-67.
16. Yegutkin GG. Enzymes involved in metabolism of extracellular nucleotides and nucleosides: functional implications and measurement of activities. Crit Rev Biochem Mol Biol 2014;49:473-97.
17. Fredholm BB, Ijzerman AP, Jacobson KA, Klotz KN, Linden J. International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol Rev 2001;53:527-52.
18. Borea PA, Gessi S, Merighi S, Varani K. Adenosine as a multi-signalling guardian angel in human diseases: when, where and how does it exert its protective effects? Trends Pharmacol Sci 2016;37:419-34.
19. Sitaraman SV, Merlin D, Wang L, Wong M, Gewirtz AT, Si-Tahar M, Madara JL. Neutrophil-epithelial crosstalk at the intestinal lumenal surface mediated by reciprocal secretion of adenosine and IL-6. J Clin Invest 2001;107:861-9.
20. Lynge J, Schulte G, Nordsborg N, Fredholm BB, Hellsten Y. Adenosine A 2B receptors modulate cAMP levels and induce CREB but not ERK1/2 and p38 phosphorylation in rat skeletal muscle cells. Biochem Biophys Res Commun 2003;307:180-7.
21. Fang Y, Olah ME. Cyclic AMP-dependent, protein kinase A-independent activation of extracellular signal-regulated kinase 1/2 following adenosine receptor stimulation in human umbilical vein endothelial cells: role of exchange protein activated by cAMP 1 (Epac1). J Pharmacol Exp Ther 2007;322:1189-200.
22. Darashchonak N, Sarisin A, Kleppa MJ, Powers RW, von Versen-Höynck F. Activation of adenosine A2B receptor impairs properties of trophoblast cells and involves mitogen-activated protein (MAP) kinase signaling. Placenta 2014;35:763-71.
23. Gao Z, Chen T, Weber MJ, Linden J. A2B adenosine and P2Y2 receptors stimulate mitogen-activated protein kinase in human embryonic kidney-293 cells. cross-talk between cyclic AMP and protein kinase c pathways. J Biol Chem 1999;274:5972-80.
24. Panjehpour M, Castro M, Klotz KN. Human breast cancer cell line MDA-MB-231 expresses endogenous A2B adenosine receptors mediating a Ca2+ signal. Br J Pharmacol 2005;145:211-8.
25. Schulte G, Fredholm BB. Signalling from adenosine receptors to mitogen-activated protein kinases. Cell Signal 2003;15:813-27.
26. Fredholm BB. Adenosine, an endogenous distress signal, modulates tissue damage and repair. Cell Death Differ 2007;14:1315-23.
27. Antonioli L, Blandizzi C, Pacher P, Haskó G. Immunity, inflammation and cancer: a leading role for adenosine. Nat Rev Cancer 2013;13:842-57.
28. Ohta A, Sitkovsky M. Role of G-protein-coupled adenosine receptors in downregulation of inflammation and protection from tissue damage. Nature 2001;414:916-20.
29. Haskó G, Linden J, Cronstein B, Pacher P. Adenosine receptors: therapeutic aspects for inflammatory and immune diseases. Nat Rev Drug Discov 2008;7:759-70.
30. Morello S, Sorrentino R, Pinto A. Adenosine A2a receptor agonists as regulators of inflammation: pharmacology and therapeutic opportunities. J Receptor Ligand Channel Res 2009:211-7.
31. Ferrari D, Malavasi F, Antonioli L. A purinergic trail for metastases. Trends Pharmacol Sci 2017;38:277-90.
32. St. Hilaire C, Carroll SH, Chen H, Ravid K. Mechanisms of induction of adenosine receptor genes and its functional significance. J Cell Physiol 2009;218:35-44.
33. Allard B, Longhi MS, Robson SC, Stagg J. The ectonucleotidases CD39 and CD73: Novel checkpoint inhibitor targets. Immunol Rev 2017;276:121-44.
34. Raskovalova T, Lokshin A, Huang X, Su Y, Mandic M, Zarour HM, Jackson EK, Gorelik E. Inhibition of cytokine production and cytotoxic activity of human antimelanoma specific CD8+ and CD4+ T lymphocytes by adenosine-protein kinase A type I signaling. Cancer Res 2007;67:5949-56.
35. Zarek PE, Huang CT, Lutz ER, Kowalski J, Horton MR, Linden J, Drake CG, Powell JD. A2A receptor signaling promotes peripheral tolerance by inducing T-cell anergy and the generation of adaptive regulatory T cells. Blood 2008;111:251-9.
36. Lappas CM, Rieger JM, Linden J. A2A adenosine receptor induction inhibits IFN-gamma production in murine CD4+ T cells. J Immunol 2005;174:1073-80.
37. Sevigny CP, Li L, Awad AS, Huang L, McDuffie M, Linden J, Lobo PI, Okusa MD. Activation of adenosine A2A receptors attenuates allograft rejection and alloantigen recognition. J Immunol 2007;178:4240-9.
38. Panther E, Corinti S, Idzko M, Herouy Y, Napp M, la Sala A, Girolomoni G, Norgauer J. Adenosine affects expression of membrane molecules, cytokine and chemokine release, and the T-cell stimulatory capacity of human dendritic cells. Blood 2003;101:3985-90.
39. Cekic C, Day YJ, Sag D, Linden J. Myeloid expression of adenosine A2A receptor suppresses T and NK cell responses in the solid tumor microenvironment. Cancer Res 2014;74:7250-9.
40. Morello S, Pinto A, Blandizzi C, Antonioli L. Myeloid cells in the tumor microenvironment: role of adenosine. Oncoimmunology 2015;5:e1108515.
41. Ohta A, Sitkovsky M. Extracellular adenosine-mediated modulation of regulatory T cells. Front Immunol 2014;5:304.
42. Stagg J, Divisekera U, McLaughlin N, Sharkey J, Pommey S, Denoyer D, Dwyer KM, Smyth MJ. Anti-CD73 antibody therapy inhibits breast tumor growth and metastasis. Proc Natl Acad Sci U S A 2010;107:1547-52.
43. Jin D, Fan J, Wang L, Thompson LF, Liu A, Daniel BJ, Shin T, Curiel TJ, Zhang B. CD73 on tumor cells impairs antitumor T-cell responses: a novel mechanism of tumor-induced immune suppression. Cancer Res 2010;70:2245-55.
44. Wang L, Fan J, Thompson LF, Zhang Y, Shin T, Curiel TJ, Zhang B. CD73 has distinct roles in non-hematopoietic and hematopoietic cells to promote tumor growth in mice. J Clin Invest 2011;121:2371-82.
45. Forte G, Sorrentino R, Montinaro A, Luciano A, Adcock IM, Maiolino P, Arra C, Cicala C, Pinto A, Morello S. Inhibition of CD73 improves B cell-mediated anti-tumor immunity in a mouse model of melanoma. J Immunol 2012;189:2226-33.
46. Loi S, Pommey S, Haibe-Kains B, Beavis PA, Darcy PK, Smyth MJ, Stagg J. CD73 promotes anthracycline resistance and poor prognosis in triple negative breast cancer. Proc Natl Acad Sci U S A 2013;110:11091-6.
47. Yegutkin GG, Marttila-Ichihara F, Karikoski M, Niemelä J, Laurila JP, Elima K, Jalkanen S, Salmi M. Altered purinergic signaling in CD73-deficient mice inhibits tumor progression. Eur J Immunol 2011;41:1231-41.
48. Quezada C, Garrido W, Oyarzún C, Fernández K, Segura R, Melo R, Casanello P, Sobrevia L, San Martín R. 5'-ectonucleotidase mediates multiple-drug resistance in glioblastoma multiforme cells. J Cell Physiol 2013;228:602-8.
49. Leclerc BG, Charlebois R, Chouinard G, Allard B, Pommey S, Saad F, Stagg J. CD73 expression is an independent prognostic factor in prostate cancer. Clin Cancer Res 2016;22:158-66.
50. Turcotte M, Spring K, Pommey S, Chouinard G, Cousineau I, George J, Chen GM, Gendoo DM, Haibe-Kains B, Karn T, Rahimi K, Le Page C, Provencher D, Mes-Masson AM, Stagg J. CD73 is associated with poor prognosis in high-grade serous ovarian cancer. Cancer Res 2015;75:4494-503.
51. Serra S, Horenstein AL, Vaisitti T, Brusa D, Rossi D, Laurenti L, D'Arena G, Coscia M, Tripodo C, Inghirami G, Robson SC, Gaidano G, Malavasi F, Deaglio S. CD73-generated extracellular adenosine in chronic lymphocytic leukemia creates local conditions counteracting drug-induced cell death. Blood 2011;118:6141-52.
52. Wennerberg E, Kawashima N, Demaria S. Adenosine regulates radiation therapy-induced anti-tumor immunity. J ImmunoTher Cancer 2015;3:378.
53. Allard B, Pommey S, Smyth MJ, Stagg J. Targeting CD73 enhances the antitumor activity of anti-PD-1 and anti-CTLA-4 mAbs. Clin Cancer Res 2013;19:5626-35.
54. Beavis PA, Divisekera U, Paget C, Chow MT, John LB, Devaud C, Dwyer K, Stagg J, Smyth MJ, Darcy PK. Blockade of A2A receptors potently suppresses the metastasis of CD73+ tumors. Proc Natl Acad Sci U S A 2013;110:14711-6.
55. Iannone R, Miele L, Maiolino P, Pinto A, Morello S. Adenosine limits the therapeutic effectiveness of anti-CTLA4 mAb in a mouse melanoma model. Am J Cancer Res 2014;4:172-81.
56. Mittal D, Young A, Stannard K, Yong M, Teng MW, Allard B, Stagg J, Smyth MJ. Antimetastatic effects of blocking PD-1 and the adenosine A2A receptor. Cancer Res 2014;74:3652-8.
57. Mittal D, Sinha D, Barkauskas D, Young A, Kalimutho M, Stannard K, Caramia F, Haibe-Kains B, Stagg J, Khanna KK, Loi S, Smyth MJ. Adenosine 2B receptor expression on cancer cells promotes metastasis. Cancer Res 2016;76:4372-82.
58. Bastid J, Cottalorda-Regairaz A, Alberici G, Bonnefoy N, Eliaou JF, Bensussan A. ENTPD1/CD39 is a promising therapeutic target in oncology. Oncogene 2013;32:1743-51.
59. Fishman P, Bar-Yehuda S, Ohana G, Barer F, Ochaion A, Erlanger A, Madi L. An agonist to the A3 adenosine receptor inhibits colon carcinoma growth in mice via modulation of GSK-3 beta and NF-kappa B. Oncogene 2004;23:2465-71.
60. Cohen S, Stemmer SM, Zozulya G, Ochaion A, Patoka R, Barer F, Bar-Yehuda S, Rath-Wolfson L, Jacobson KA, Fishman P. CF102 an A3 adenosine receptor agonist mediates anti-tumor and anti-inflammatory effects in the liver. J Cell Physiol 2011;226:2438-47.
61. Jajoo S, Mukherjea D, Watabe K, Ramkumar V. Adenosine A(3) receptor suppresses prostate cancer metastasis by inhibiting NADPH oxidase activity. Neoplasia 2009;11:1132-45.
62. Morello S, Sorrentino R, Montinaro A, Luciano A, Maiolino P, Ngkelo A, Arra C, Adcock IM, Pinto A. NK1.1 cells and CD8 T cells mediate the antitumor activity of Cl-IB-MECA in a mouse melanoma model. Neoplasia 2011;13:365-73.
63. Montinaro A, Forte G, Sorrentino R, Luciano A, Palma G, Arra C, Adcock IM, Pinto A, Morello S. Adoptive immunotherapy with Cl-IB-MECA-treated CD8+ T cells reduces melanoma growth in mice. PLoS One 2012;7:e45401.
64. Silverman MH, Strand V, Markovits D, Nahir M, Reitblat T, Molad Y, Rosner I, Rozenbaum M, Mader R, Adawi M, Caspi D, Tishler M, Langevitz P, Rubinow A, Friedman J, Green L, Tanay A, Ochaion A, Cohen S, Kerns WD, Cohn I, Fishman-Furman S, Farbstein M, Yehuda SB, Fishman P. Clinical evidence for utilization of the A3 adenosine receptor as a target to treat rheumatoid arthritis: data from a phase II clinical trial. J Rheumatol 2008;35:41-8.
65. Fishman P, Bar-Yehuda S, Liang BT, Jacobson KA. Pharmacological and therapeutic effects of A3 adenosine receptor agonists. Drug Discov Today 2012;17:359-66.
66. Avni I, Garzozi HJ, Barequet IS, Segev F, Varssano D, Sartani G, Chetrit N, Bakshi E, Zadok D, Tomkins O, Litvin G, Jacobson KA, Fishman S, Harpaz Z, Farbstein M, Yehuda SB, Silverman MH, Kerns WD, Bristol DR, Cohn I, Fishman P. Treatment of dry eye syndrome with orally administered CF101: data from a phase 2 clinical trial. Ophthalmology 2010;117:1287-93.
67. David M, Gospodinov DK, Gheorghe N, Mateev GS, Rusinova MV, Hristakieva E, Solovastru LG, Patel RV, Giurcaneanu C, Hitova MC, Purcaru AI, Horia B, Tsingov II, Yankova RK, Kadurina MI, Ramon M, Rotaru M, Simionescu O, Benea V, Demerdjieva ZV, Cosgarea MR, Morariu HS, Michael Z, Cristodor P, Nica C, Silverman MH, Bristol DR, Harpaz Z, Farbstein M, Cohen S, Fishman P. Treatment of plaque-type psoriasis with oral CF101: data from a Phase II/III multicenter, randomized, controlled trial. J Drugs Dermatol 2016;15:931-8.
68. Yang D, Koupenova M, McCrann DJ, Kopeikina KJ, Kagan HM, Schreiber BM, Ravid K. The A2b adenosine receptor protects against vascular injury. Proc Natl Acad Sci U S A 2008;105:792-6.
69. Sun CX, Zhong H, Mohsenin A, Morschl E, Chunn JL, Molina JG, Belardinelli L, Zeng D, Blackburn MR. Role of A2B adenosine receptor signaling in adenosine-dependent pulmonary inflammation and injury. J Clin Invest 2006;116:2173-82.
70. Eckle T, Faigle M, Grenz A, Laucher S, Thompson LF, Eltzschig HK. A2B adenosine receptor dampens hypoxia-induced vascular leak. Blood 2008;111:2024-35.
71. Grenz A, Osswald H, Eckle T, Yang D, Zhang H, Tran ZV, Klingel K, Ravid K, Eltzschig HK. The reno-vascular A2B adenosine receptor protects the kidney from ischemia. PLoS Med 2008;5:e137.
72. Ryzhov S, Novitskiy SV, Zaynagetdinov R, Goldstein AE, Carbone DP, Biaggioni I, Dikov MM, Feoktistov I. Host A(2B) adenosine receptors promote carcinoma growth. Neoplasia 2008;10:987-95.
73. Cagnina RE, Ramos SI, Marshall MA, Wang G, Frazier CR, Linden J. Adenosine A2B receptors are highly expressed on murine type II alveolar epithelial cells. Am J Physiol Lung Cell Mol Physiol 2009;297:L467-74.
74. Yang D, Zhang Y, Nguyen HG, Koupenova M, Chauhan AK, Makitalo M, Jones MR, St Hilaire C, Seldin DC, Toselli P, Lamperti E, Schreiber BM, Gavras H, Wagner DD, Ravid K. The A2B adenosine receptor protects against inflammation and excessive vascular adhesion. J Clin Invest 2006;116:1913-23.
75. Casadó V, Casillas T, Mallol J, Canela EI, Lluis C, Franco R. The adenosine receptors present on the plasma membrane of chromaffin cells are of the A2b subtype. J Neurochem 1992;59:425-31.
76. Peakman MC, Hill SJ. Adenosine A2B-receptor-mediated cyclic AMP accumulation in primary rat astrocytes. Br J Pharmacol 1994;111:191-8.
77. Corset V, Nguyen-Ba-Charvet KT, Forcet C, Moyse E, Chédotal A, Mehlen P. Netrin-1-mediated axon outgrowth and cAMP production requires interaction with adenosine A2b receptor. Nature 2000;407:747-50.
78. Nishida K, Dohi Y, Yamanaka Y, Miyata A, Tsukamoto K, Yabu M, Ohishi A, Nagasawa K. Expression of adenosine A2b receptor in rat type II and III taste cells. Histochem Cell Biol 2014;141:499-506.
79. Hua X, Kovarova M, Chason KD, Nguyen M, Koller BH, Tilley SL. Enhanced mast cell activation in mice deficient in the A2b adenosine receptor. J Exp Med 2007;204:117-28.
80. Novitskiy SV, Ryzhov S, Zaynagetdinov R, Goldstein AE, Huang Y, Tikhomirov OY, Blackburn MR, Biaggioni I, Carbone DP, Feoktistov I, Dikov MM. Adenosine receptors in regulation of dendritic cell differentiation and function. Blood 2008;112:1822-31.
81. Mirabet M, Herrera C, Cordero OJ, Mallol J, Lluis C, Franco R. Expression of A2B adenosine receptors in human lymphocytes: their role in T cell activation. J Cell Sci 1999;112:491-502.
82. Fredholm BB, Irenius E, Kull B, Schulte G. Comparison of the potency of adenosine as an agonist at human adenosine receptors expressed in Chinese hamster ovary cells. Biochem Pharmacol 2001;61:443-8. in Chinese
83. Eltzschig HK, Ibla JC, Furuta GT, Leonard MO, Jacobson KA, Enjyoji K, Robson SC, Colgan SP. Coordinated adenine nucleotide phosphohydrolysis and nucleoside signaling in posthypoxic endothelium: role of ectonucleotidases and adenosine A2B receptors. J Exp Med 2003;198:783-96.
84. Zhao P, Li XG, Yang M, Shao Q, Wang D, Liu S, Song H, Song B, Zhang Y, Qu X. Hypoxia suppresses the production of MMP-9 by human monocyte-derived dendritic cells and requires activation of adenosine receptor A2b via cAMP/PKA signaling pathway. Mol Immunol 2008;45:2187-95.
85. Feoktistov I, Ryzhov S, Zhong H, Goldstein AE, Matafonov A, Zeng D, Biaggioni I. Hypoxia modulates adenosine receptors in human endothelial and smooth muscle cells toward an A2B angiogenic phenotype. Hypertension 2004;44:649-54.
86. Zhong H, Belardinelli L, Maa T, Zeng D. Synergy between A2B adenosine receptors and hypoxia in activating human lung fibroblasts. Am J Respir Cell Mol Biol 2005;32:2-8.
87. Kong T, Westerman KA, Faigle M, Eltzschig HK, Colgan SP. HIF-dependent induction of adenosine A2B receptor in hypoxia. FASEB J 2006;20:2242-50.
88. Xaus J, Mirabet M, Lloberas J, Soler C, Lluis C, Franco R, Celada A. IFN-gamma up-regulates the A2B adenosine receptor expression in macrophages: a mechanism of macrophage deactivation. J Immunol 1999;162:3607-14.
89. Németh ZH, Leibovich SJ, Deitch EA, Vizi ES, Szabó C, Hasko G. cDNA microarray analysis reveals a nuclear factor-kappaB-independent regulation of macrophage function by adenosine. J Pharmacol Exp Ther 2003;306:1042-9.
90. St Hilaire C, Koupenova M, Carroll SH, Smith BD, Ravid K. TNF-alpha upregulates the A2B adenosine receptor gene: the role of NAD(P)H oxidase 4. Biochem Biophys Res Commun 2008;375:292-6.
91. Nguyen DK, Montesinos MC, Williams AJ, Kelly M, Cronstein BN. Th1 cytokines regulate adenosine receptors and their downstream signaling elements in human microvascular endothelial cells. J Immunol 2003;171:3991-8.
92. Schingnitz U, Hartmann K, Macmanus CF, Eckle T, Zug S, Colgan SP, Eltzschig HK. Signaling through the A2B adenosine receptor dampens endotoxin-induced acute lung injury. J Immunol 2010;184:5271-9.
93. Kolachala VL, Wang L, Obertone TS, Prasad M, Yan Y, Dalmasso G, Gewirtz AT, Merlin D, Sitaraman SV. Adenosine 2B receptor expression is post-transcriptionally regulated by microRNA. J Biol Chem 2010;285:18184-90.
94. Nakatsukasa H, Tsukimoto M, Harada H, Kojima S. Adenosine A2B receptor antagonist suppresses differentiation to regulatory T cells without suppressing activation of T cells. Biochem Biophys Res Commun 2011;409:114-9.
95. Warabi M, Kitagawa M, Hirokawa K. Loss of MHC class II expression is associated with a decrease of tumor-infiltrating T cells and an increase of metastatic potential of colorectal cancer: immunohistological and histopathological analyses as compared with normal colonic mucosa and adenomas. Pathol Res Pract 2000;196:807-15.
96. Shi B, Vinyals A, Alia P, Broceno C, Chen F, Adrover M, Gelpi C, Price JE, Fabra A. Differential expression of MHC class II molecules in highly metastatic breast cancer cells is mediated by the regulation of the CIITA transcription Implication of CIITA in tumor and metastasis development. Int J Biochem Cell Biol 2006;38:544-62.
97. Fang M, Xia J, Wu X, Kong H, Wang H, Xie W, Xu Y. Adenosine signaling inhibits CIITA-mediated MHC class II transactivation in lung fibroblast cells. Eur J Immunol 2013;43:2162-73.
98. Xia J, Fang M, Wu X, Yang Y, Yu L, Xu H, Kong H, Tan Q, Wang H, Xie W, Xu Y. A2b adenosine signaling represses CIITA transcription via an epigenetic mechanism in vascular smooth muscle cells. Biochim Biophys Acta 2015;1849:665-76.
99. Ben Addi A, Lefort A, Hua X, Libert F, Communi D, Ledent C, Macours P, Tilley SL, Boeynaems JM, Robaye B. Modulation of murine dendritic cell function by adenine nucleotides and adenosine: involvement of the A(2B) receptor. Eur J Immunol 2008;38:1610-20.
100. Wilson JM, Ross WG, Agbai ON, Frazier R, Figler RA, Rieger J, Linden J, Ernst PB. The A2B adenosine receptor impairs the maturation and immunogenicity of dendritic cells. J Immunol 2009;182:4616-23.
101. Xaus J, Valledor AF, Cardó M, Marquès L, Beleta J, Palacios JM, Celada A. Adenosine inhibits macrophage colony-stimulating factor-dependent proliferation of macrophages through the induction of p27kip-1 expression. J Immunol 1999;163:4140-9.
102. Ryzhov S, Novitskiy SV, Goldstein AE, Biktasova A, Blackburn MR, Biaggioni I, Dikov MM, Feoktistov I. Adenosinergic regulation of the expansion and immunosuppressive activity of CD11b+Gr1+ cells. J Immunol 2011;187:6120-9.
103. Gabrilovich DI, Ostrand-Rosenberg S, Bronte V. Coordinated regulation of myeloid cells by tumours. Nat Rev Immunol 2012;12:253-68.
104. Cekic C, Sag D, Li Y, Theodorescu D, Strieter RM, Linden J. Adenosine A2B receptor blockade slows growth of bladder and breast tumors. J Immunol 2012;188:198-205.
105. Iannone R, Miele L, Maiolino P, Pinto A, Morello S. Blockade of A2b adenosine receptor reduces tumor growth and immune suppression mediated by myeloid-derived suppressor cells in a mouse model of melanoma. Neoplasia 2013;15:1400-9.
106. Morello S, Miele L. Targeting the adenosine A2b receptor in the tumor microenvironment overcomes local immunosuppression by myeloid-derived suppressor cells. Oncoimmunology 2014;3:e27989.
107. Wei Q, Costanzi S, Balasubramanian R, Gao ZG, Jacobson KA. A2B adenosine receptor blockade inhibits growth of prostate cancer cells. Purinergic Signal 2013;9:271-80.
108. Vecchio EA, Tan CY, Gregory KJ, Christopoulos A, White PJ, May LT. Ligand-independent adenosine A2B receptor constitutive activity as a promoter of prostate cancer cell proliferation. J Pharmacol Exp Ther 2016;357:36-44.
109. Kasama H, Sakamoto Y, Kasamatsu A, Okamoto A, Koyama T, Minakawa Y, Ogawara K, Yokoe H, Shiiba M, Tanzawa H, Uzawa K. Adenosine A2b receptor promotes progression of human oral cancer. BMC Cancer 2015;15:563.
110. Gessi S, Merighi S, Varani K, Cattabriga E, Benini A, Mirandola P, Leung E, Mac Lennan S, Feo C, Baraldi S, Borea PA. Adenosine receptors in colon carcinoma tissues and colon tumoral cell lines: focus on the A(3) adenosine subtype. J Cell Physiol 2007;211:826-36.
111. Ma DF, Kondo T, Nakazawa T, Niu DF, Mochizuki K, Kawasaki T, Yamane T, Katoh R. Hypoxia-inducible adenosine A2B receptor modulates proliferation of colon carcinoma cells. Hum Pathol 2010;41:1550-7.
112. Wang P, Guo X, Zong W, Song B, Liu G, He S. MicroRNA-128b suppresses tumor growth and promotes apoptosis by targeting A2bR in gastric cancer. Biochem Biophys Res Commun 2015;467:798-804.
113. Long JS, Schoonen PM, Graczyk D, O'Prey J, Ryan KM. p73 engages A2B receptor signalling to prime cancer cells to chemotherapy-induced death. Oncogene 2015;34:5152-62.
114. Hajiahmadi S, Panjehpour M, Aghaei M, Shabani M. Activation of A2b adenosine receptor regulates ovarian cancer cell growth: involvement of Bax/Bcl-2 and caspase-3. Biochem Cell Biol 2015;93:321-9.
115. Daniele S, Zappelli E, Natali L, Martini C, Trincavelli ML. Modulation of A1 and A2B adenosine receptor activity: a new strategy to sensitise glioblastoma stem cells to chemotherapy. Cell Death Dis 2014;5:e1539.
116. Desmet CJ, Gallenne T, Prieur A, Reyal F, Visser NL, Wittner BS, Smit MA, Geiger TR, Laoukili J, Iskit S, Rodenko B, Zwart W, Evers B, Horlings H, Ajouaou A, Zevenhoven J, van Vliet M, Ramaswamy S, Wessels LF, Peeper DS. Identification of a pharmacologically tractable Fra-1/ADORA2B axis promoting breast cancer metastasis. Proc Natl Acad Sci U S A 2013;110:5139-44.
117. Przybylski M. A review of the current research on the role of bFGF and VEGF in angiogenesis. J Wound Care 2009;18:516-9.
118. Feoktistov I, Goldstein AE, Ryzhov S, Zeng D, Belardinelli L, Voyno-Yasenetskaya T, Biaggioni I. Differential expression of adenosine receptors in human endothelial cells: role of A2B receptors in angiogenic factor regulation. Circ Res 2002;90:531-8.
119. Feoktistov I, Ryzhov S, Zhong H, Goldstein AE, Matafonov A, Zeng D, Biaggioni I. Hypoxia modulates adenosine receptors in human endothelial and smooth muscle cells toward an A2B angiogenic phenotype. Hypertension 2004;44:649-54.
120. Merighi S, Simioni C, Gessi S, Varani K, Mirandola P, Tabrizi MA, Baraldi PG, Borea PA. A(2B) and A(3) adenosine receptors modulate vascular endothelial growth factor and interleukin-8 expression in human melanoma cells treated with etoposide and doxorubicin. Neoplasia 2009;11:1064-73.
121. Zeng D, Maa T, Wang U, Feoktistov I, Biaggioni I, Belardinelli L. Expression and function of A 2B adenosine receptors in the U87MG tumor cells. Drug Develop Res 2003;58:405-11.
122. Sorrentino C, Miele L, Porta A, Pinto A, Morello S. Myeloid-derived suppressor cells contribute to A2B adenosine receptor-induced VEGF production and angiogenesis in a mouse melanoma model. Oncotarget 2015;6:27478-89.
123. Murdoch C, Muthana M, Coffelt SB, Lewis CE. The role of myeloid cells in the promotion of tumour angiogenesis. Nat Rev Cancer 2008;8:618-31.
124. Allard B, Turcotte M, Spring K, Pommey S, Royal I, Stagg J. Anti-CD73 therapy impairs tumor angiogenesis. Int J Cancer 2014;134:1466-73.
125. Sorrentino C, Miele L, Porta A, Pinto A, Morello S. Activation of the A2B adenosine receptor in B16 melanomas induces CXCL12 expression in FAP-positive tumor stromal cells, enhancing tumor progression. Oncotarget 2016;7:64274-88.
126. Orimo A, Gupta PB, Sgroi DC, Arenzana-Seisdedos F, Delaunay T, Naeem R, Carey VJ, Richardson AL, Weinberg RA. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell 2005;121:335-48.
Cite This Article
Export citation file: BibTeX | RIS
OAE Style
Sorrentino C, Morello S. Role of adenosine in tumor progression: focus on A2B receptor as potential therapeutic target. J Cancer Metastasis Treat 2017;3:127-38. http://dx.doi.org/10.20517/2394-4722.2017.29
AMA Style
Sorrentino C, Morello S. Role of adenosine in tumor progression: focus on A2B receptor as potential therapeutic target. Journal of Cancer Metastasis and Treatment. 2017; 3: 127-38. http://dx.doi.org/10.20517/2394-4722.2017.29
Chicago/Turabian Style
Sorrentino, Claudia, Silvana Morello. 2017. "Role of adenosine in tumor progression: focus on A2B receptor as potential therapeutic target" Journal of Cancer Metastasis and Treatment. 3: 127-38. http://dx.doi.org/10.20517/2394-4722.2017.29
ACS Style
Sorrentino, C.; Morello S. Role of adenosine in tumor progression: focus on A2B receptor as potential therapeutic target. J. Cancer. Metastasis. Treat. 2017, 3, 127-38. http://dx.doi.org/10.20517/2394-4722.2017.29
About This Article
Copyright

Data & Comments
Data


 Cite This Article 13 clicks
Cite This Article 13 clicks

 Like This Article 34
likes
Like This Article 34
likes






Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.