
Bacteria in cancer therapy: beyond immunostimulation

Abstract
Currently, conventional therapies in cancer are improving; chemotherapy, radiotherapy and surgery have increased survival significantly. New therapies have arisen with the same goal; immunotherapy has appeared as a promising option in the fight against cancer stimulating the immune system by inducing innate and adaptive responses. These responses include release of pro-inflammatory cytokines, making the immune system capable to eliminate or protect against multiple tumors. Nowadays, many of these therapies are being used in clinical settings, such as checkpoint inhibitors, monoclonal anti cytotoxic T-Lymphocyte associated protein 4 (CTL-4) and programmed death protein 1 (PD1), with inspiring results; however, they may decrease immunotolerance, limiting their use. At the same time, chemotherapy works by passive transport across the cell membrane, limiting its capacity to penetrate in tumor cells. For these reasons, bacteria employment represents one of the best candidates for cancer treatment. They can surpass these barriers with their selective colonization which also has an oncolytic effect by increasing proliferation and immunostimulation in the tumor environment. Attenuated strains, such as Mycobacterium bovis, Clostridium, Salmonella typhimirium and Listeria monocytogenes have been studied showing promising results in experimental models. However, their application in clinical trials has shown the need to maximize their therapeutic effect. Genetic engineering and synthetic biology are necessary to prove the scope that this novel approach has against cancer due to implications of cancer therapy and public health.
Keywords
Introduction
At present, cancer has one of the highest morbidity and mortality rates worldwide, nationwide and statewide[1,2]. It comes from the growth of uncontrolled and invasive malignant cells with DNA mutations capable of producing multiple diseases[3]. Most of these malignant neoplasms have the same etiopathogenesis. However, the diversity on the anatomic location, histologic origin, immunologic characteristics and intrinsic spreading capacity (intertumoral heterogeneity)[4], and different genomic alterations inside the same tumor (intratumoral heterogeneity)[5] have shown the need for specific biomarkers and individualized therapy to improve patient prognosis.
Currently, conventional therapy such as surgery, chemotherapy, radiotherapy, or mixed therapy have increased survival rates worldwide against cancer in different subtypes[6,7]. However, these practices produce many adverse effects and have shown a limited tumor penetrance[8]. The role of the immune system has been studied in order to find a therapeutic approach with equivalent therapeutic potency and controlled damage to healthy tissue; which gave rise to immunotherapy as a novel treatment[9].
Nowadays, many of these therapies are being used in clinical settings, including the checkpoint inhibitors monoclonal antibodies anti cytotoxic T-Lymphocyte associated protein 4 (CTL-4) and programmed death protein 1 (PD1). They have been shown to increase survival in patients with metastatic melanoma[10] but their mechanism of action decreases immunotolerance with systemic administration. The latter may cause autoimmune adverse effects, limiting its use only for specific patients[11]. In the last few decades, experimental studies and clinical trials have been aimed to assess bacteria therapeutic functions[12-15]. Bacteria selective replication within the tumor microenvironment gives them antitumor effect and minimizes systemic adverse effects. On the other hand, expression of multiple ligands, immunostimulants, cytokines and tumor antigens can be achieved through gene manipulation to increase the therapeutic effect against specific tumors.
Cancer causes many physical and psychological effects to the patients and their families, but it also increases state expenditures. For these reasons, evaluation of these novel therapies in clinical settings has great importance. This review brings the basic science principles in genetics, immunology, and microbiology that gave rise to this therapeutic approach, in addition to its latest experimental and clinical advances.
The beginning: genetic and immunologic basis in carcinogenesis
Carcinogenesis begins as a result of multiple genomic alterations within a cell. They come from a prolonged exposure to different mutagens[16], adverse epigenetic factors[17], as well as chronic infections[18]. These alterations increase proliferation and affect cell cycle through gene functioning[19], in proto oncogenes[20] and tumor suppressor genes[21], causing different mutations[22]. They ultimately modify the cell physiology making a mutated cell capable to generate its own mitogenic signals, resist against growth inhibitory signals, and acquires its own blood vessels. In advanced stages it can even invade and metastasize[3].
The role of the immune system in tumor surveillance comes from the response to multiple oncogenic viruses and other infecting agents that can induce a chronic inflammatory environment leading to carcinogenesis[23,24]. Identification of tumor-specific antigens (TSA) induces an immune response on carcinogenesis at an early stage[24]. Tumor cells generate multiple modified surface proteins, decreasing immunologic tolerance as carcinogenesis progresses, and many TSA are expressed[25]. The immune system can recognize and eliminate abnormal cells, in a continuous and bidirectional pathway between innate and adaptive immunity, which is called “Immunosurveillance”[26]. Natural killer (NK) cells[27] and cytotoxic T lymphocytes (T-CD8+) are the main mediators in this process[28]; macrophages associated to tumors, dendritic cells (DC), naïve T cells, aβT-cell receptor (TCR)-expressing T cells, γδT-cells and regulatory T cells (T-reg) FOXP3+ also participate in the immune response towards the tumor. They interact with tumor cells, while some act inhibiting their growth and others stimulate it, composing the tumor microenvironment[29].
NK cells are considered the main part of the innate immunity against tumors. They recognize and eliminate neoplastic cells effectively[30,31]; but are not confined to the innate immune system. They also act with the adaptive immunity by working as T-lymphocyte response modulators[32]. Damage associated molecule patterns (DAMPs) are released from tumor cell elimination mediated by NK cells[33], increasing DC maturation[34] and presentation to T-CD8+ lymphocytes on major histocompatibility complex (MHC)-1 molecules[35]. Once activated, NK cells and T-CD8+ lymphocytes induce activation, proliferation and recruiting of other cells to the tumor site[36]. This is achieved through the release of cytokines such as interferon gamma (IFN-γ), granulocyte and macrophage colonies stimulating factor (GM-CSF) and tumor necrosis factor (TNF)[37]. IFN-γ carries important functions such as direct inhibition of tumor growth, macrophage activation, and increases Th1 expression among T-CD4+ lymphocytes. This represents their major role in modulating cellular response against tumors[38]. T-CD8+ lymphocytes require the expression of tumor antigens on MHC-1 molecules and co-stimulatory signals in the tumor site in order to function appropriately[39].
Cancer may become clinically detectable in advanced stages explained by the mechanisms in which tumor cells evade immune surveillance[40]. This theory started with the “immunoediting” process[41], where the immune system works inversely: making an immunosuppressed environment that favors tumor growth. This process is composed by three phases: elimination, equilibrium, and escape [Figure 1], being the elimination phase a homologous mechanism from those seen in immunosurveillance[42].
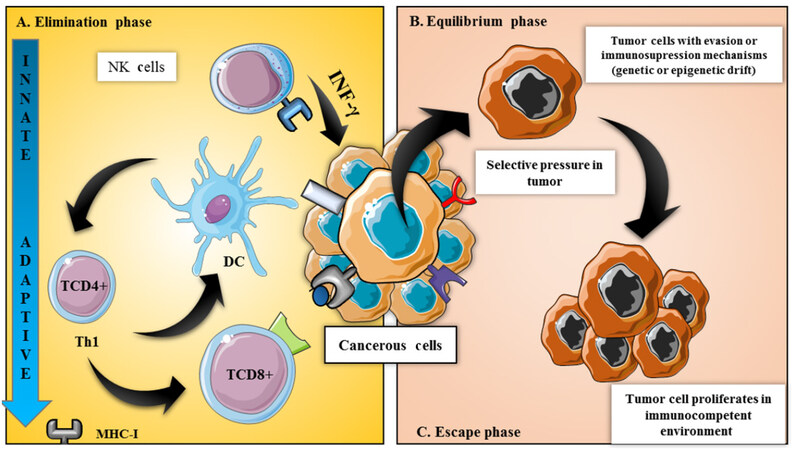
Figure 1. Immunosurveillance vs. immunoediting: key points in regulation of immune system in tumor progression/regression. Immunoediting comprises 3 phases: (A) elimination: when the tumorcells begins to proliferate, an inflammatory response is induced by the injured tissue. This causes the migration of cells from the immune system, orchestrating the innate immune response; (B) the equilibrium phase: in which this continuous process produces a selective pressure in these cells that can cause genetic or epigenetic rearrangement, causing certain cells to evade these immunological effector mechanisms; (C) the escape phase, when cells that have evaded these mechanisms also gained uncontrolled growth ability. DC: dendritic cells; MHC: major histocompatibility complex; NK: natural killers
Once the tumor cell has escaped the elimination phase, it enters the equilibrium phase[43]. This phases consists in the destruction of cells expressing tumor antigens in their MHC-1 molecules by T-CD8+[44]. Following this, less immunogenic cellular clones will be immunoselected and more aggressive tumor cells will grow, directing them to the escape phase[43]. Although evasion of the immune system is not an isolated event; it also includes an immunological adaptation process. During this process an immunosuppressed microenvironment comes with recruitment T-reg FOXP3+[45] and release of immunomodulatory molecules such as transforming growth factor β (TGFβ), prostaglandin E2 (PGE2), indoleamin 2,3 dioxygenase (IDO), adenosine, and interleukin-10 (IL 10); also with decreased expression of co-stimulatory proteins and increased expression of inhibitor molecules such as CTL-4/CD28 and PD-1/PD-L1, called checkpoints[46].
The discovery of these processes has led to research looking for novel immunologic therapies against cancer[47]. Most of this therapy approaches have been dedicated to increase active or passive immune responses. Others have tried to modify tumor cells to increase recognition by the immune system[48]. Despite of this, only few immunotherapies have achieved a response strong enough to be clinically effective[49]. For these reasons, using bacteria to potentiate response has become a promising strategy.
Incoming bacteriology: enemies or allies?
Chronic infection with biological agents represents a risk factor associated with cancer, with viral agents leading in this field[50]. Bacteria have been associated with cancer because of their effect on cell cycle, and their capability to evade the immune system and cause immunosuppression through chronic infections[51-53]. Bacterial infections stimulate phagocyte activity and increase oxidative stress on neighboring cells. The latter causes the release of reactive oxygen (ROS) and nitrogen (RNS) species such as peroxynitrite (ONOO-), reactive hydroxyl group (OH-) and other free radicals that damage cell membranes and DNA, affecting enzymatic activity and gene expression[54]. Among DNA alterations mediated by oxidative stress, the most common includes the formation of 8-oxoguanine and/or 8-2’-desoxyguanosine. These modified nucleotides are caused by deregulated and repetitive metabolism, and lead to mutagenesis by inhibiting or enabling expression of altered genes[55]. Chronic inflammation is considered carcinogenic[56] by activation and preservation of nuclear factor ĸB (NF-κB) [Figure 2] which modulates gene expression related to cell cycle[57,58], apoptosis[59,60], proinflammatory cytokines, angiogenic processes[58], invasion and metastasis[61,62].
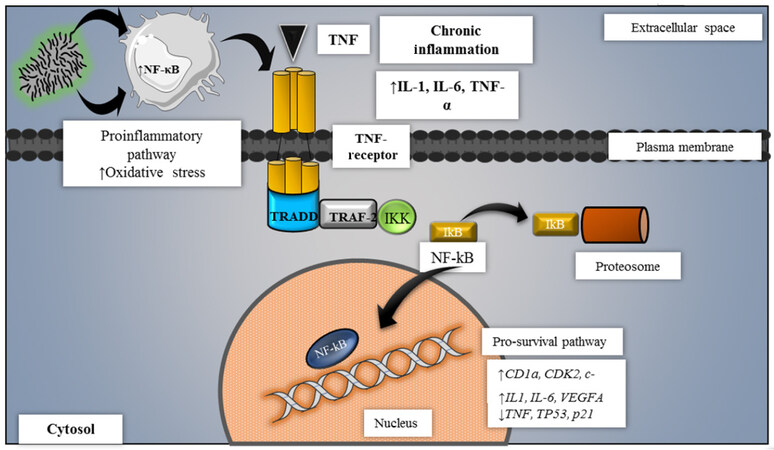
Figure 2. Carcinogenesis molecular mechanisms associated to chronic inflammation. Chronic inflammatory cascade is carcinogenic by the activation of the NF-kB pathway. This leads to the degradation of such proteins, allowing that NF-kB enter the nucleus to mediate the transcription of specific cell cycle-related genes while genes responsible for apoptosis are downregulated. IKK: I kappa B kinase; NF-kB: nuclear factor kappa B; TNF: tumor necrosis factor; TRADD: tumor necrosis factor receptor type 1-associated DEATH domain protein; TRAF: TNF receptor-associated factor; IkB: I-kappa-B
Infectious agents can act directly on the genome of their carrier and promote carcinogenesis by inactivation of tumor suppressor genes or mitotic stimulation. For example, chronic infections with Helicobacter pylori (H. pylori) carrying CagA positive virulence factor, causes mutations on p53 protein and adenomatous polyposis coli (APC) tumor suppressor genes; it can also induce loss of deleted in colorectal carcinoma (DCC) gene and microsatellite instability[52]. Cases of infection by Mycobacterium tuberculosis affect tissue structure, generating a fibrotic scar that will probably increase the risk of carcinogenesis by blocking the lymphatic flow that decreases activated leucocyte depuration and increases risk for metastatic deposits. Mycobacterium tuberculosis can also modulate tumor immunity together with the frequent co-infection with the human immunodeficiency virus (HIV), promoting survival of the bacillus and inhibiting INF-γ secretion with increase in TNF-α secretion[63].
Epidemiologic studies support a relationship between bacteria and cancer. Salmonella tiphy chronic carrier state is related to gallbladder cancer[64], Streptoccocus bovis, found in bacteremia complications and infective endocarditis, is related to colorectal tumors[53], H. pylori, known by its relationship with gastric adenocarcinoma, is also related to esophageal cancer[52], and Chlamydia pneumoniae has been considered as an etiological factor in patients with lung cancer[51].
Learning from Coley’s toxin
Human carcinogenesis is not related to all bacteria[65]. Some bacterial properties work through mechanisms that stimulate the immune system and are capable to potentiate defenses against malignancy[66]. Bacteria’s role against cancer was recognized in the 19th century, when an American oncologist, Dr. William Coley observed tumor regression in patients with acute bacterial infections[67]. After this observation, he decided to administer inactivated Streptococcus pyogenes and Serratia marcescens[68] - in a mixture he called Coley’s toxin - to a patient with an inoperable sarcoma, inducing tumor regression and curing the patient[69-71]. Furthermore, it was used in cases with carcinomas, lymphomas, melanomas and myelomas, having significant results[72].
The Coley’s toxin mechanism of action became a key finding for immunotherapy[73]. It is composed of gram-negative bacterial endotoxin (Serratia marcescens), a lipopolysaccharide released from the bacterial cell membrane that was considered a prototype for pathogen associated molecular patterns (PAMPs). This compound induces the secretion of TNF, IL-2, INF-α and IL-12[74] from the immune system. Being IL-12 the most important in both innate and adaptive immunities since it stimulates T-CD4+ Th1 cells development, and increases NK/NKT and TCD8+ lymphocytes pathway mechanisms[75].
These pathways require Preexistent Immunization in order to gain antineoplastic activity. This comes from expression of IL-12 receptors only on activated T cells[76], explaining its major effectiveness in patients with previously sensitized T cells[76]. On the other hand, bacterial intrinsic properties could also be used against tumors, such as Streptokinase from Streptococcus pyogenes cases, an enzyme considered as one of the active agents in Coley’s toxin. This enzyme has anti-angiogenic effects, suppressing new vessel formation and decreasing tumor growth and invasion[77]. Despite the fact that some clinical trials have shown effectiveness with this therapy, others have not shown any success, presenting multiple reasons for treatment failures[78]. High doses of IL-12 used as support treatment with other cytokines have produced an immunologic response with high toxicity and its employment has been cancelled[74].
Bacteria supporting the fight against cancer: a crossroad for genetics, immunology and microbiology
Aiming for the perfect bacterium
Limited penetration in tumor tissue is considered a challenge for conventional therapy. This happens to chemotherapy and other specific biological therapies. They all depend on passive transport of the molecules into the tumor, limiting their efficacy and increasing their risk for toxicity[8]. On the other hand, bacterial therapy works through mechanisms against cancer that cannot be achieved with standard conventional methods, becoming a great prospect[79].
The main issue with therapeutic uses of microorganisms against cancer in the 19th century was the adverse effects associated to immunity, such as fever, septic shock, and death[80,81]. Development in genetic engineering has led to use genetically modified bacteria- decreasing their pathogenicity- as cancer therapy[82]. Their accessible genome manipulation make bacteria the best candidates among other microorganisms[83]. Giving them the ability to enter cancerous tissue[82,84], selecting tumor cells following specific chemical signals in their microenvironment[85,86] and acting as vectors for molecule transportation[87,88] assuming the fact they can be controlled from outside [Figure 3][89-92].
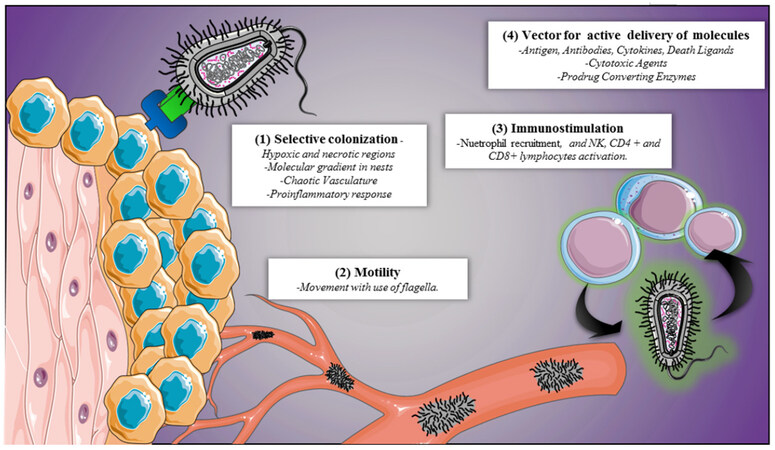
Figure 3. Bacterial therapeutic mechanisms. Multiple bacterial features that can be used to make novel therapies against cancer. NK: natural killer
The “artificial medical bacteria” also have a role in the diagnostic process (detecting molecules or tumor markers related to certain diseases), therapeutic decision making (detection of chemical stimuli and production of therapeutic agents) and most importantly, can be controlled[93]. Synthetic biology has been used to design and build biologic machineries based on vehicles. Bacteria compounds integrated on genes, proteins and molecules coming from multiple origins can affect their security and therapeutic effect[94]. Systemic administration of these compounds would be better. Less concentration would be needed and multiple agents could be made without requiring neither formulation nor purification processes to amplify their effect[93].
Selective colonization in cancerous tissue
Blood supply in cancerous tissue is insufficient, which results in acidity, deprivation of nutrients and presence of hypoxic areas[95]. Hypoxia is more associated with expression of malignant phenotypes characterized by genomic instability, angiogenesis and metastatic qualities[96]; leading to new approaches against this feature[97-100]. Strict anaerobic bacteria with spore germination qualities, such as Clostridium, cannot proliferate in highly oxygenated environments, restricting their colonization to hypoxic and necrotic regions of the tumor[101]. This is affected by tumor morphology, with central necrotic areas and well perfused cells in the periphery, allowing anaerobic bacteria to proliferate in the center. Following this, the immune system gets activated and makes a peripheral ring of immune cells prepared to eradicate the tumor completely[102].
Facultative anaerobic bacteria such as Salmonella act differently. They are capable to identify and penetrate into tumors by detecting chemotactic factors including molecular gradients of serine, aspartate, and ribose[86,103]. Necrotic cancer cells release these compounds after being exposed to hypoxia for prolonged periods of time[103]. A strain of obligate anaerobic Salmonella has been associated with antitumor features by replacing the asd gene with recombinant technology. Making the gene expressed only with hypoxia-inducible promoters[104] to maximize selective colonization. Selective colonization consists of the bacteria’s ability to be confined to the tumor chaotic vasculature[105]. An increase in cytokines production such as TNF-α is observed in response to primary colonization of bacteria that leads to a secondary colonization[106]. In addition to this, auxotrophic microorganisms-capable to grow in environments with nutrients produced only in tumor nests- have been synthesized with mutations generated from null alleles lacking biological capability[107,108].
Recently, bacterial motility has shown to be critical in tumor colonization. Many bacteria have flagella that work with consumption of energy[109]. Bacteria use this feature to migrate and stay for longer periods of time on places distal to tumor vasculature, in contrast to passive transport with chemotherapy[83]. In addition to this, differences between diffusion and pressure gradients limit movement of molecules by passive transport and most of this happens on poorly perfused tumor areas[105].
Immunostimulation in tumor microenvironment
There is no bacterium capable of completely inhibiting tumor growth just through colonization[110]. However, it represents an important prospect for cancer treatment as an immunostimulator or as a vector for therapeutic components that can be released inside a tumor[111,112][Table 1].
Pre-clinical studies for evaluation of molecular antitumor effects made by genetic engineering bacteria
| Bacterium | Molecule | Most relevant results | Reference |
|---|---|---|---|
| Salmonella typhimurium | |||
| VNP20009 | CCL21 | Increased intratumoral production INFg, CXCL9 and CXCL1 | Loeffler et al.[87] |
| VNP20009 | LIGHT (TNFSF14) | Prominent reduction in tumor growth was observed. Evidenced with an inflammatory infiltrate (B lymphocytes, CD4+,CD8+ in models treated with this bacterium | Loeffler et al.[113] |
| VNP20009 | IL-18 | Inhibition of tumor growth was observed. Evidenced with a leukocytic infiltrate (especially NK cells) and increased secretion of INF-g, TNF-a, IL-1b and GM-CSF | Loeffler et al.[114] |
| VNP20009 | FASL | Significant reduction of tumor size was observed in primary tumors and lung metastases, increasing neutrophil recruitment | Loeffler et al.[115] |
| VNP20009 | TRAIL | TRAIL expression increased tumor cells apoptosis dependent on caspase 3 and 8 | Ganai et al.[116] |
| S. choleraesuis | Endostatine | Inhibition of tumor growth was observed in 40%-70%. Evidenced with a decrease in intratumoral microvasculature, VEGF expression and increase in T CD8+ lymphocyte recruitment | Lee et al.[117] |
| S. choleraesuis | Thrombospondin | Selective colonization was observed in a 1000:1 to 10000:1 ratio with respect to liver and spleen. Evidenced with inhibition of tumor growth and increase in survival by angiogenic effects | Lee et al.[118] |
| Nula phoP/phoQ LH430 | RNAi-STAT3 | RNAi inhibited significantly tumor growth, the number of metastatic lessions decreased, increased survival rate in animal models | Zhang et al.[119] |
| S. typhiTy21 | VEGFR-2 | Vaccination for this molecule showed inhibition of tumor growth, decreased metastasis growth and prevented new spontaneous metástasis, increasing survival rate in models | Niethammer et al.[120] |
| aroA SL7207 | PSA-CtxB* | This vaccine administration conjugated with Salmonella showed protective effects by reducing tumor size in 8-14 days since its inoculation. This mechanism depends on T CD8+ lymphocyte activity and a prototype of the E. coli Hemolysin secretion system | Fensterle et al.[121] |
| Clostridium | |||
| C. beijerinckii | NR | Nitroreductase activity increased in vitro antitumor activity of CB in 1954, by a factor of 22 | Lemmon et al.[122] |
| C. beijerinckii | Citosine deaminase | Tumor cells sensitivity to 5-fluorocytosine increased by 500 times | Fox et al.[123] |
| C. sporogenes | IL-12 | Increased selective secretion of INF-g with effects on tumor growth, without signs of toxicity | Zhang et al.[124] |
| C. novyi-NT | AC anti-HIFa | A heterologous gene transfer was satisfactory in this bacterium. Showing increased antibody secretion (with adhesion capacity and specificity) | Groot et al.[125] |
| Listeria monocytogenes | |||
| Lm-LLO-E7 | HPV16-E7* | This therapy induced regression in 75% of tumors expressing E7 antigen. This response depends on TCD4+ and TCD8+ lymphocytes and INFg secretion | Gunn et al.[126] |
| ADXS31–164 | HER-2/neu (Human)* | An increase in TCD8/Tregs ratio was observed with this therapy. It also prevented more breast tumor formation and delayed more metastasis growth than other vaccines based on this bacterium | Shahabi et al.[127] |
| LM-LLO-Mage-b/2nd | MAGE-b* | The most effective vaccine for breast tumors, decreasing number of metastasis by 96%, correlating to a strong CD8+ lymphocytic response in spleen after restimulation with antigen use | Kim et al.[128] |
| Lm-LLO-HMW-MAA-C | HMW-MAA | This therapy immunization prevented tumor growth not only in models that expressed the antigen, but in melanoma, renal carcinoma and breast carcinoma. TCD4+ and TCD8+ lymphocytes were needed to achieve this | Maciag et al.[129] |
The main theory for this approach comes from the bacterial intrinsic ability to immunostimulate after colonizing tumor tissue. They can proliferate inside the tumour where an increased activity of the immune system has been observed. Neutrophils, T CD8+ and CD4+ cells recruitment, cytokine and chemokine release, potentiate immune response with no effect on the surrounding healthy tissue[130]. This approach has shown better results than conventional therapy since it can affect healthy and cancerous tissue altogether.
Bacteria have one of the largest genomes that exist. They can express multiple therapeutic transgenes and increase immune activity with cytokines and tumor antigens presentation[131]. They can transfer those genes to eukaryotic cells and get expressed or repressed[132,133]. Systemic administration of cytokines such as IL-2, IL-8, and CCL21Al may show certain limitations related to their short half-life and adverse effects[134]. Their manufacture is highly expensive and they lack tumor orientation, which may cause severe systemic inflammatory reactions restricting their clinical use[134]. In contrast, gene modified bacteria are manufactured with low expenses, can be directed to specific tumor tissue, and may be easily eradicated with antibiotics[114,135]. Bacteria in situ cytokine production may benefit those with difficult DNA recombination methods and/or protein instability in production and purification. To achieve oncolytic activity genes are introduced to increase cytokine expression and promote tumor regression[113].
Antitumor activity can be achieved without significant toxicity and related to inflammatory cell infiltration such as granulocytes, T lymphocytes and NK cells. Induction of intratumor production of cytokines[114,136,137], including IL-18, is important to enhance cytokine production in T lymphocytes and NK cells, to increase MHC-1 expression, and to favor differentiation of Th1 CD4+ cells; leading to an immune response mediated by NK cells, macrophages, and T CD8+ cells[114,138].
Bacteria induce expression of ligands in cancerous cells with antitumor activity. For example, the FAS ligand (FASL), member of TNF family, enhances chemotaxis and IL-23 production from dendritic cells with T cell proliferation[115]. TNF related to apoptosis inducing ligand (TRAIL) protein expression has been achieved in models with breast cancer[116], gastric cancer[139] and melanoma by employment of controlled bacteria[140].
An interesting fact about cancer prognosis is the advanced stage by the time it is diagnosed, decreasing patient survival. Therefore, bacteria have been employed to work as vaccine vectors. These vaccines would increase tumor antigen expression on cancerous cells. Among these: prostatic specific antigen (PSA) in prostate cancer[121], C-rapidly accelerated fibrosarcoma in pulmonary adenoma[141], and alpha-fetoprotein (α-FP) for hepatocellular carcinoma[142]. They can work by inducing an adaptive immune response to protect against these tumors. APCs recognition of these antigens is followed by a specific T-CD8+ cell proliferation with immunologic memory, in contrast to systemic administration of antibodies or adoptive T cell leading to loss of immunotolerance and healthy tissues affected[143].
Other use as therapeutic vectors
Expression of hemolytic toxins could be achieved in tumors resistant to conventional therapy. Cytolisin A (Clya) with E. coli K-12 use[144] is an example of these. Transcription factors could be induced in cases with S. Typhimurioum JRG5356 where genes for HlyE activation are expressed so pore-forming cytolysins are made by activating the FF+20 promoter[85]. On the other hand, inhibition of angiogenetic processes with TSP-1 or endostatin genes could be used to decrease capillary density and reduce expression of vascular endothelial growth factors (VEGF)[117,118].
Lastly, protein repression could also be induced using RNA interference (RNAi). Bacteria carrying plasmids such as pSi-Stat3 are capable of changing specific portions of DNA and increase expression of small interference RNA (siRNA)[145] or short hairpin RNA (shRNA). All of these cause degradation of specific mRNA sequences leading to a dysfunctional tumor gene expression[146,147].
Potential of bacteria utilization in oncology
Research on bacteria employment against malignant tumors in human subjects is expanding in diagnostics (for their selective colonization and external control) and therapeutics (for their antitumor effect). The next sections will discuss experimental and clinical evidence supporting bacteria utilization against cancer.
Bacteria utilization as cancer diagnostic method and to monitor therapeutic efficacy
Bacteria utilization is not limited to the therapeutic scope but also to diagnostic methods. Developments in genetic engineering have shown expression of bacterial genes that can be detected and monitored externally by fluoroscopy[148], magnetic resonance imaging (MRI)[90] and positron emission tomography (PET)[149] scan. These genes can code for light-emitting proteins, such as luciferase and green fluorescent protein (GFP), making them observable in real time under low light image processing; and also under micro-to-micro fluorescent microscopes[150]. E. coli bacterium remains as a prototype carrying PLITE201 plasmid that codes for luxCDABE protein[151] giving luminescent features. This bacterium also carries the pMW211 plasmid that codes for dsred protein[152] making cancerous cells recognizable in their exact localization by turning them luminescent without any invasive approach. Salmonella typhimyrium and Vibrio cholerae remain under study for their utilization in colon and breast cancer diagnosis, respectively[153].
MRI is routinely used for tumor diagnosis and treatment evaluation. Magneto spirillum is a bacterium employed in this radiologic study. It consists of a microaerophilic microorganism with magnetic properties on its magnetosome which contains magnetic crystals formed mainly of magnetite (Fe3O4) covered by a lipid bilayer membrane[154]. Experiments with AMB-1 strains of this bacterium have shown positive contrast features in T1-enhanced imaging when they were cultured under iron deprived (FeCl3) conditions[90]. Contrast was intensified with expression of MagA gene. This gene codes for an iron transporter that gets positively regulated in presence of low iron concentrations[155]. In vitro experiments with colon carcinoma models in HT-29 human subjects did not show any evidence of toxicity and tumor necrosis was observed on both histologic slides and MRI[156]. Employment of these magnetic features implies new advances with clinical use potential.
Regarding PET scans, other modified strains of E. coli: E. coli Nissle (EcN) 1917[152] along with pyrimidine nucleotide analogs have been considered for diagnosis of breast tumors. Increased local accumulation of radio-isotopes has shown a positive correlation with the number of bacteria containing radioactive drugs. These bacteria selectively colonized tumors, making them detectable via PET. Other bacterium employed for these studies was Salmonella VNP20009-TK. The latter has had similar results with a positive correlation between intratumor bacteria and fialuridine sequestration (FIAU), a radio-marked nucleoside analog used for tumor identification[157].
Other bacteria diagnostic features can be used in oncology. This includes their employment as probiotics in cancer screening[158]. Use of EcN with modified genetic circuits enhances detection of focal metastasis in urine samples. Two principles were applied: first, bacteria produced a luminous signal that can be detected through imaging techniques; and second, LacZ enzymatic activity on a substrate composed of luciferine and galactose (LuGal) results in luciferine traveling into the circulatory system and serving as a colorimetric indicator with fluorescent or luminescent features; these features could be detected in urine samples[158].
Current perspective in bacteria based therapy in medicine
Recently, bacterial strains with therapeutic characteristics against cancer have been discovered. Mycobacterium bovis (BCG) is considered a prototype. It is an obligate anaerobic, acid-alcohol-resistant, facultative intracellular and non-motile bacterium that has been employed in the past for tuberculosis vaccine manufacturing[159]. For more than 30 years it has been utilized in bladder cancer patients as immunotherapy. A decrease in tumor recurrence has been observed with this therapy along with its well tolerated adverse effects[160]. Even though the first-choice treatment for non-muscle invasive bladder cancer (NMIBC) is still transurethral resection for bladder tumor (TURBT), a high recurrence (50%-70%) and progression rates (10%-20%) after two years have been observed with this procedure[161]. In these cases, intravesical instillation with BCG is one of the main therapeutic options, decreasing long term appearance of distant metastasis[162]; also an increase in global survival in 5 years with long term maintenance of this therapy has been observed[163]. However, its long term use may have adverse effects such as drug induced cystitis, hematuria, and systemic toxicity[164]. Despite of this, BCG is still considered the standard treatment for NMIBC after transurethral resection for bladder tumor has been performed in patients with intermediate and high risk of progression or recurrence[12].
The mechanism behind these benefits has not been clarified, but the antitumoral effects of BCG are considered to come from the immune response[165,166]. Once urothelial cells or macrophages internalize the bacillus, they induce an immune response with secretion of TNF-α, IL-6, IL-10, INF-γ, FEC-GM, CC and expression of CXC chemokine receptors[167-169]; this stimulates recruitment of neutrophils, macrophages, T-CD4+ cells and increases expression of MHC-I, MHC-II and IL-2 receptors[170-172]. TRAIL is one of the main mediators in bacteria based therapy. This ligand appears to be upregulated in response to INF-γ, causing urothelial cell death[173,174]. BCG remains as reference for novel cancer therapies in development such as vaccines, and also for nonbacterial therapies, having similar efficacy and reliability (NCT02010203). Next sections discuss the most important bacteria used for these goals, going from their experimental research to current clinical evidence [Table 2].
Clinical evidence evaluating the safety, tolerance, adverse and therapeutic effects of bacteria against cancer
| Reference | Bacterium/compound | Metodology | Clinical phase | Results |
|---|---|---|---|---|
| Nemunaitis et al.[175] | Samonella TAPET-CD | Open clinical trial that included 3 patients with solid and/or metastatic tumors, 5-FU sensistive, without any response to coventional therapies. With intratumor administration of bacteria | Phase I | A favorable response was observed in 2 patients at their injection site. Bacterial CD dependent conversion of 5-FC to 5-FU. Presented adverse effects not related to therapy |
| Toso et al.[110] | Salmonella VNP20009. | Open clincal trial that included 24 patients with metastatic melanoma and one pacient with renal cell carcinoma to assess safety, tolerability and clinical response | Phase I | From the 25 patients treated with VNP20009, none experimented an objective tumor regression. Dose-limiting toxicity was associated to TNF-α and IL1-β secretion, despite the majority of adverse effects showed reversibility |
| Schmitz-Winnenthal et al.[176] | S. typhiTy21/ Anti-VEGFR-2 (VXM01) | Ramdomized, double-blind clinical trial to assess safety, tolerability, and clinical and immunologic responses in 45 patients with locally advanced stage IV pancreatic cancer | Phase I | Treatment was well toleraed in all applied doses. No dose-limiting toxicity was found. There was an effector T lymphocyte dependent response and a decrease in tumor perfussion in patients with preexisting immunologic memory |
| Roberts et al.[13] | Clostridium novyi-NT | Clinical trial including 1 patient with retroperitoneal leiomyosarcoma and received intratumoral administration of spores in a metastatic lession on shoulder | Phase I | Extensive tumor destruction was observed, compatible with necrosis. By day 4 after administration, biopsy showed absence of viable tumor cells. By day 55, presented with a pathologic fracture. Therapy improved his quality of life |
| Maciag et al.[185] | Lm-LLO-E7 | Non ramdomized clinical trial to assess safety of the therapy in 15 patients with advanced stage cevrical cancer, refractary or recurrent | Phase I | Ll patients presented adverse effects, including severe (grade 3) in 6 patients (40%). At the end of the study, 2 patients died, 5 developed disease progression, 7 showed stable disease and partial tumor response was observed in one patient |
| Le et al.[177] | ANZ-100/CRS-207 | Open multicentric clinical trial to assess safety and induction of immune system in two groups: 1) ANZ-100 = 9 patients with colorectal cancer (6), pancreatic cancer (2), and melanoma (1). 2) CRS-207 = 17 patients with pancreatic cancer (7), mesothelioma (5), lung cancer (3) and ovarian cancer (2) | Phase I | In both groups, therapy was well tolerated with self-limited adverse effects. In group 1, no dose-limiting toxicity was found with ANZ-100 administration, and was related to NK cell (CD38) activation and increase in MCP-1, MIP-1β and INFγ secretion. In group 2, CRS-207 was well tolerated. The majority of observed adverse effects were grade 2. Like IN group 1, an increase in proinflammatory cytokines was observed. CRS-207 induced a specific response dependent on T cells towards mesotheline and listeriolysin-O |
| Le et al.[178] | CRS-207 | Ramdomized multicentric clinical trial to assess safety and clinical response in 90 patients with stage IV pancreatic adenocarcinoma and administration of Cy/GVAX+CRS-207 (A) in contrast to Cy/GVAX only (B) | Phase II | The mean follow-up was 3.4 months. The global survival rate was higher in patients treated with Cy/GVAX+CRS-207 (n = 61) than those treated with GVAX/Cy only (n = 29) (HR: 0.59; IC 95%: 0.36-0.97, P = 0.02). Nevertheless, increase in T CD8+ lymphocytic specific response to mesothelin was associated to a higher global survival rate, independent on treatment group |
Clostridium: heading to tumor specificity
Necrosis and hypoxia in tumor tissues make them resistant to conventional therapies[106], therefore, research on Clostridium began because of its natural anaerobic features[101]. In regards to utilization of this gram-positive, obligate anaerobe, spore forming bacteria to developa therapy against cancer in tumors with necrosis associated to bad prognosis[179-181], Clostridium novyi is one of the most studied. After a deletion of its α-toxine gene, Clostridium novyi-NT becomes capable to colonize selectively; in addition to diminished adverse effects because of its decreased exotoxin production[82]. It was used in experimental models with colorectal cancer, renal carcinoma[99], gliomas[182], and sarcomas[13] to observe its selective colonization, immune cell infiltration, and cytokine release leading to tumor tissue necrosis[125]. Phase I clinical trials were initiated on one patient with retroperitoneal leiomyosarcoma presenting multiple metastasis and refractoriness to conventional therapy. After intratumoral application-preferred over systemic administration to decrease adverse effects - the tumor located in this patient right shoulder regressed with an extensive necrotic area; and medically managed adverse effects[13,183]. Further clinical trials are currently in patient recruitment (NCT01924689).
Genetic therapy was employed to increase the oncolytic effects of this strain and presented promising results[122,184,185]. Clostridium sporogenes[13] was utilized because of its tumor directed features. Genes derived from E. coli serve for nitroreductase (NR) and cytosine deaminase (CD) codification[185,186]. These enzymes metabolize cytotoxic drugs inside tumors, having in vivo antitumoral effects.
Clostridium spores have low immunogenicity and can colonize multiple organs after systemic administration[187]. However, once they germinate, they induce an inflammatory response with infiltration of immune cells with oncolytic effects[99]. These strains have been employed in genetic engineering as vectors for cytokines secretion such as TNF-α[188], IL-12[124] and IL-2[189], achieving high concentrations inside tumor tissue without systemic toxicity. Also C. novyi-NT and C. sporogenes increase secretion of specific antibodies against hypoxia inducing factor-1 (HIF-1), main component observed in hypoxia response regulation inside tumors[125].
Salmonella: multi-use bacterium
Salmonella enterica serivar tyhimurium (S. typhimurium) is one of the most studied bacterium for its adaptative qualities leading to new strains with bacterial engineering showing antitumor activity[119]. In the beginnings of the 21st century, phase I clinical trials were conducted to show their efficacy with gene modification via deletion in msbB and purI genes. The msbB gene is required for lipid A synthesis and its deletion was made to reduce TNF-α related toxicity, preventing septic shock[190]. On the other hand, by deletion of purI gene, the bacterium became able to colonize tumors more selectively. All of this made the strains depend on purine external sources for survival restricting their growth to areas with substantial cell renewal[191]. Tumor tissues with their purine rich activities would be perfect regions for their selective colonization[191]. Salmonella typhimurium VNP20009 is one of the main strains in experimental studies originated from this theory.
This study results showed the maximum tolerated dose of this bacteria, its toxicity limit dose, and adverse effects by increasing production of proinflammatory cytokines. The observed adverse effects included thrombocytopenia, anemia, persistent bacteremia, hyperbilirubinemia, nausea, vomit, elevated alkaline phosphatase and hypophosphatemia. However, tumor colonization was detected only in 3 patients, and no tumor regression was observed[110]. Despite the fact that the study did not show promising results, it was the start line for prospect studies to find doses that could be adjusted for efficiency and tumor localization and for other therapeutic features.
In order to increase this bacterium therapeutic effect, a study was initiated to use them as vectors in tumor gene therapy[192]. A pilot study was performed with an attenuated and gene modified Salmonella strain with expression of E. coli CD, called suicide prodrug-activating enzyme[193]. These genes were integrated in VNP20009 chromosome through Donnenberg and Karper method resulting in TAPET-CD strain[175]. The mechanism of action of this enzyme consists in conversion of 5-fluorocytosine (5-FC, antifungal agent with limited cytotoxicity) to 5-fluorouracil (5-FU, cytotoxic antimetabolite capable of producing cellular apoptosis)[193]. No promising results were obtained 2 out of 3 treated patients did not present any tumor regression but an improvement of their disease was observed[175].
Another example from S. typhimurium is strains would be A1-R, which currently is on preclinical studies against different cancerous tissues such as prostate[14], pancreatic[194,195], glioma[196], colorectal[197], and ovary[198]. S. typhimurium A1-R colonization seems to be more selective and effective than other strains and less toxic than VNP20009 strain. It also has safer systemic administration than C. novyNT[199]. Therefore, clinical trials for this strain are coming. A1-R is a gene modified strain, auxotrophic for leucine and arginine by nitroguanidine mutagenesis (NTG)-preventing healthy tissue invasion. It was utilized in animal models with prostatic cancerous cells PC3 and also in humans showing tumor regression, inhibition and prevention of cancer[14].
Bacteria therapeutic use has been confirmed in cancer models with stem cell characteristics. This represents the only method capable to reduce in vivo tumor sizes in relation to chemotherapy (5-FU in monotherapy, cisplatin and gemcitabine). The efficacy increased when combined with 5-FU[195]. S. typhimurium A1-R could induce cell entrance from G0/G1 to S/G2/M and reduced significant portion of cells in quiescent state, making them sensible to chemotherapy[200,201].
Other approach in gene modification of Salmonella was the study of strains to decrease or inactivate gene expression. This inactivation could be achieved with utilization of iRNA[119,202]. S. typhimurium LH340 strain was made with deletion on popP/phoQ operon required for its virulence resulting in its attenuation[203,204]. The signal transducer and activator of transcription protein-3 (STAT3) is the goal with these therapies. Confirmation of its role in immune system depression[205] and expression of target genes such as VEGF, Cyclin D1, Cyclin D2, c-Myc, p53, Bcl-XL, Bcl-2, Mcl-1 and Survivin have been observed[206]. A relationship between inhibition of these genes expression and suppression of tumor growth was found[207].
Strains with expression of iRNA for Stat3 suppressed tumor growth significantly, reduced metastasis and increased survival in experimental models with prostate[119] and hepatocellular carcinoma[202]. These tumors are usually highly vascularized and angiogenesis inhibition through plasmids required for endostatin codification (SL/pEndostatin) may increase efficacy to the novel therapy[133]. By introducing Stat3, (SL/pEndo-Si-Stat3) more antitumor effects were observed. These effects were related to angiogenesis inhibition and increase in TCD8+ lymphocyte proliferation, NK cytotoxicity and T-regs proliferation. The later came from inhibition probably by stimulation of INF-γ and TNF-α secretion with significant decrease in TGF-β concentrations[202].
In clinical settings Salmonella typhi Ty21a is one of the new therapy prospects. It was studied to find a vaccine to prevent typhoid fever[173]. The bacterium was introduced to cancer therapy strategies with the VXM01 vaccine. This is an oral vaccine made of live attenuated strains of S. typhi ty21a capable to induce a T cell response; it also contains a plasmid that codes for VEGFR2 and plays an important role in tumor angiogenesis[208]. It can also induce both humoral and cellular responses[176] observed in experimental models with melanoma, colorectal cancer and lung cancer. Suppression of primary tumor growth and metastatic lesions mediated by T-CD8+ cells activity was observed in these models[117]. In clinical settings, it was recently evaluated on 45 patients with stage IV pancreatic cancer and it showed the importance of preexisting immunologic memory for effector T cells to achieve an antiangiogenic effect[176].
Clinical trials have shown that Salmonella still lacks therapeutic efficacy and selective tumor colonization but could be considered as a multi-use bacterium for its diverse features. It can work as a vector, and a better inducer of antitumoral response because of its efficient type III secretion system[209]. Prospecting studies should be focused on this objective with specific molecules for each cancer type, getting major effectiveness.
Listeria monocytogenes: the perfect antigenic vector
Listeria monocytogenes (Lm) is a gram positive, facultative intracellular bacterium[131]. Over the last few decades multiple studies have shown that it can work cancer therapeutic agent with multiple effect or mechanisms[210]. It can be used against primary and metastatic tumors in an immune-privileged microenvironment. The latter helps its selective colonization and favors their elimination with ROS production[211]. In addition to this, Lm decreases T-regs cells and immunomodulation molecules such as TGFβ and IL10 in tumor microenvironment[212]. However, main feature of Lm consists on selectively infecting APCs favoring self-antigen and heterolog antigens processing and presentation[213]. These characteristics make Lm to be considered as a valuable immunostimulant agent.
Intracellular life cycle of Lm favors its use as an immunotherapeutic agent. Once infection has ocurred, Lm strongly activates innate immunity with the release of proinflammatory cytokines such as IL-2, IL-6, IL-12, and TNF-α; and increases expression of co-stimulant molecules in APCs surfaces leading to maturation and activation of high affinity T cells[214]. After internalization by phagocytes, Lm is capable to escape from phagolysosomes using its virulence factor called listeriolysin O (LLO)[215]. It works as a hemolysin that perforates the phagosomal membranes of the bacterium could escape into the cytosol. Once in the cytosol, they can replicate and secrete its antigens[216]. This mechanism makes antigen processing and presentation to be via both class I and II MHC molecules[217] inducing potent specific responses from both T-CD4+ and T-CD8+ cells[218].
These features of Lm have been studied with genetic engineering looking for recombinant strains capable to secrete tumor antigens[219]. They could be employed as live vectors through vaccines to potentiate cellular response and overcome immunotolerance towards certain types of cancers[131]. This could be achieved with insertion of plasmids encoding the tumor antigen[126], or by their integration in the bacterial chromosome[220]. These antigens would be expressed as chimeric proteins along with Lm virulence factors[221] such as LLO or actin assembly inducing-protein (ActA)[222]. Lm uses ActA for motility and intercellular propagation and its immunogenic features increase the immune response towards tumor antigens with poor immunogenicity[223]. These experimental studies were oriented to measure efficacy in recently developed vaccines. Among these vaccines, Lm-LLO-E7 was studied for cervical cancer models[224], Lm-her2-neu for metastatic breast cancer[211], Lm-LLO-PSA for prostate cancer[225], Lm-MPFG for hepatocellular carcinoma[226] and LM-Kras for pancreatic ductal adenocarcinoma[227] and others; all of them reporting suppression in growth and even regression[228].
Lm utilization as live vector could induce systemic disease in immunocompromised individuals limiting its use for human vaccines[221]. Different strains have been cultured with specific gene deletionsto guarantee their safety[229-232]. Among these new strains, only XFL-7 and LmΔactA/ΔplcB have been used in clinical trials. The XFL-7 strain was created with chromosomal deletion in its Prfa gene. This gene codes for an activating transcription factor needed for bacterial virulence factor expression. In order to increase its expression, a complementation of a multicopy plasmid with a heterolog gene was introduced[231]. The LmΔactA/ΔplcB strain was made with a deletion of its virulence genes ActA and inlB-used for surface proteins codification that favors cell invasion-to prevent capture from non-phagocytic cells and reduce hepatic damage[232].
The first clinical trial to assess safety with Lm administration in cancer patients utilized attenuated strains as vaccines, specially Lm-LLO-E7[126]. The latter was made from XLF-7 strains to express E7 oncoantigen from human papilloma virus serotype 16 (HPV16). This vaccine was also designed to treat cervical cancer[15], and other tumors induced by HPV16 such as oropharyngeal cancer[224]. In this open, nonrandomized, uncontrolled study, Maciag et al.[185] assessed safety and viability of Lm-LLO-E7 via intravenous administration with intervals of 21 days. Doses of 1 × 109, 3.3 × 109 or 1 × 1010 Colony-Forming Units (CFU) were administered to 15 patients with invasive cervical carcinoma in advanced stages and refractory to conventional therapy. Despite the fact that all the patients presented systemic adverse effects in the study (fever, vomit, headache, muscle aches, tachycardia, hypotension, anemia) most of them were alleviated during the first 12-h post dose, responding to symptomatic treatment whenever necessary[15].
Safety of Lm-LLO-E7 administration in humans is still under study with insertion of plasmid encoded resistance to chloramphenicol required for bacterial survival in vivo[233]. Phase II clinical trials to assess efficacy and safety in patients with oropharyngeal cancer (NCT01598792) were suspended after a patient developed systemic listeriosis following vaccination[234]. This shows the need for a new attenuation, especially for their use on immunocompromised patients.
The Listeria strain LmΔactA/ΔplcB with application of two vaccines called ANX-100 and CRS-207 has been studied[177]. ANX-100 consisted of a vector without antigen that was administered to 9 patients with colon cancer and hepatic metastasis from colon cancer and demonstrated its safety and tolerability to a dose of 1 × 108 CFU. It induced an antitumor inflammatory response. CRS-207 consisted of a modified strain to express mesothelin, which is an overexpressed antigen that is frequently found in multiple solid tumors, including mesothelioma, pancreatic adenocarcinoma, non-small cell lung carcinomas and ovarian cancer[235-237]. Phase I clinical trials in patients with these characteristics showed their efficacy and tolerability to a dose of 1 × 109 CFU[177]. Seven patients were treated during these trials. Six patients had increased survival in 15 months, showing treatment efficacy. But 3 patients with high survival rates had been treated with GVAX previously. This vaccine was designed to increase GM-CSF expression for its ability to induce cellular immunity against tumor antigens. Phase II clinical trials were performed posteriorly[178]. They evaluated the safety and efficacy of the combined treatment with GVAX and cyclophosphamide (GVAX/Cy) with CRS-207 in contrast to exclusive administration of GVAX/Cy in patients with pancreatic cancer. Reports showed a global survival rate of 6.1 months in patients treated with GVAX/Cy+CRS-207, more than patients treated with GVAX/Cy exclusively (HR: 0.59; 95% CI: 0.36-0.97, P = 0.02).
Based on these results, current research is focusing on efficacy evaluation of vaccines based on Lm attenuated strains along with other immunological or conventional therapies. Among these, combining LM-LLO-E7 with anti-PD1 antibodies[238], or using the strain as adjuvant therapy after chemotherapy against cervical cancer (NCT02853604). There was also found that combination of CRS-207 strain with an IDO1 inhibitor increases immunotherapeutic effects in ovarian and peritoneal cancer treatment (NCT02575807); which could be used as adjuvant therapy after chemotherapy for malignant pleural mesothelioma (NCT01675765).
Other bacteria under study
Research for bacteria use in cancer treatment is not limited to the cited genres. Lactococcus lactis NK34, generally used as a probiotic, showed significant antitumor activity against lung, colorectal, gastric and breast cancers on in vitro models[239]. These effects appear to be mediated by an increase in tumor expression of p21 and p53 leading to apoptosis[240,241]. Intratumor Streptoccous pyogenes was employed in pancreatic cancer models and complete tumor regression was observed and associated to cytokine release and immune cell infiltration[242]. Recently, Bacillus subtilis and Bifidobacterium infantis are being included in preclinical studies to find more evidence supporting bacteria as life-saving prospects[243-245].
Conclusion
The main advantage of bacterial therapy is its selective colonization in tumor tissue decreasing its toxicity. This direct oncolytic effect resides on proliferation and immunostimulation that take place in cancerous tissues. Despite lacking significant effects in initial models and multiple adverse effects, it has overcome these barriers. Development in genetic engineering has led to better therapeutic effects and the reinforcement of therapies with molecules such as cytokines, tumor antigens, drug metabolizing enzymes, death receptors, and even RNA interference. Promising results have been observed with these therapies during clinical trials. Research is beginning to determine their use as main, or supportive therapy in contrast to conventional therapy against cancer. Their toxicity, antitumor effect, and their long half-life represent critical variables to consider in future research protocols and clinical trials. However, microorganisms versatility remains a feature that may show encouraging results in the future [Table 3] with significant improvements in cancer diagnosis and treatment.
Current clinical trials to evaluate bacteria use in cancer treatment
| Bacterium | Indicaction | Clinical phase | NCT identification |
|---|---|---|---|
| Clostridium novyi-NT | Patients with malignant solid tumors refractory to treatment | Phase I | NCT01924689 |
| Lm-LLO-E7 | Patients with grade 2 cervical intraepithelial neoplasia | Phase II | NCT01116245 |
| Patients with non-small cell lung carcinoma, HPV positive | Phase II | NCT02531854 | |
| Patients with anorectal carcinoma | Phase II | NCT02399813 | |
| Patients with HPV positive oropharyngeal cancer | Phase II | NCT02002182 | |
| Patients with high risk of locally advanced cervical cancer | Phase III | NCT02853604 | |
| CRS-207 | Adults with previously treated pancreatic adenocarcinoma | Phase II | NCT02004262 |
| Patients with malignant pleural mesothelioma | Phase I | NCT01675765 | |
| Patients with ovarian or peritoneal cancer | Phase I/II | NCT02575807 | |
| ADU-623 | Patients with astrocytic tumors | Phase I | NCT01967758 |
| ADXS31-142 | Patients with prostate cancer | Phase I/II | NCT02325557 |
Declarations
Authors’ contributionsArticle conception and design: Olivar LC, Lameda V, Torres W
Acquisition and analysis of bibliographic information: Torres W, Martinez MS, Lameda V, Olivar LC, Navarro C, Fuenmayor J, Pérez A, Rojas M
Drafting of the manuscript: Torres W, Martinez MS, Lameda V, Olivar LC, Navarro C, Fuenmayor J, Pérez A, Rojas M, Rojas J
Critical revision and final approval: Bermúdez V, Velasco M, Mindiola A, Rojas J
Financial support and sponsorshipNone.
Conflict of interestsThe authors have no conflict of interest.
Patient consentNot applicable.
Ethics approvalNot applicable.
Copyright© The Author(s) 2018.
REFERENCES
1. Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D, Bray F. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer 2015;136:E359-86.
2. Ministery of Health. Bolivarian Republic of Venezuela. Anuary of Mortality 2013. Available from: http://www.ovsalud.org/descargas/publicaciones/documentos-oficiales/Anuario-Mortalidad-2013.pdf [Last accessed on 19 Jan 2018].
3. Hainaut P, Plymoth A. Targeting the hallmarks of cancer: towards a rational approach to next-generation cancer therapy. Curr Opin Oncol 2013;25:50-1.
4. Burrell R, McGranahan N, Bartek J, Swanton C. The causes and consequences of genetic heterogeneity in cancer evolution. Nature 2013;501:338-45.
5. Fisher R, Pusztai L, Swanton C. Cancer heterogeneity: implications for targeted therapeutics. Br J Cancer 2013;108:479-85.
6. Spratt D, Pei X, Yamada J, Kollmeier M, Cox B, Zelefsky M. Long-term survival and toxicity in patients treated with high-dose intensity modulated radiation therapy for localized prostate cancer. Int J Radiat Oncol Biol Phys 2013;85:686-92.
7. Senn H. Gallen consensus 2013: optimizing and personalizing primary curative therapy of breast cancer worldwide. Breast Care 2013;8:101.
8. Khawar I, Kim J, Kuh H. Improving drug delivery to solid tumors: Priming the tumor microenvironment. J Control Release 2015;201:78-89.
9. Nagarsheth N, Wicha M, Zou W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat Rev Immunol 2017;17:559-72.
10. Larkin J, Chiarion-Sileni V, Gonzalez R, Grob J, Cowey C, Lao C, Schadendorf D, Dummer R, Smylie M, Rutkowski P, Ferrucci PF, Hill A, Wagstaff J, Carlino MS, Haanen JB, Maio M, Marquez-Rodas I, McArthur GA, Ascierto PA, Long GV, Callahan MK, Postow MA, Grossmann K, Sznol M, Dreno B, Bastholt L, Yang A, Rollin LM, Horak C, Hodi FS, Wolchok JD. Combined nivolumab and ipilimumab or monotherapy in untreated melanoma. N Engl J Med 2015;373:23-34.
11. Wolchok J, Chiarion-Sileni V, Gonzalez R, Rutkowski P, Grob J, Cowey C, Lao C, Wagstaff J, Schadendorf D, Ferrucci P, Smylie M, Dummer R, Hill A, Hogg D, Haanen J, Carlino M, Bechter O, Maio M, Marquez-Rodas I, Guidoboni M, McArthur G, Lebbé C, Ascierto P, Long G, Cebon J, Sosman J, Postow M, Callahan M, Walker D, Rollin L, Bhore R, Hodi F, Larkin J. Overall survival with combined Nivolumab and Ipilimumab in advanced melanoma. N Engl J Med 2017;377:345-56.
12. Babjuk M, Böhle A, Burger M, Capoun O, Cohen D, Compérat E, Hernández V, Kaasinen E, Palou J, Rouprêt M, van Rhijn BWG, Shariat SF, Soukup V, Sylvester RJ, Zigeuner R. EAU guidelines on non-muscle-invasive urothelial carcinoma of the bladder: update 2016. Eur Urol 2017;71:447-61.
13. Roberts N, Zhang L, Janku F, Collins A, Bai R, Staedtke V, Rusk AW, Tung D, Miller M, Roix J, Khanna KV, Murthy R, Benjamin RS, Helgason T, Szvalb AD, Bird JE, Roy-Chowdhuri S, Zhang HH, Qiao Y, Karim B, McDaniel J, Elpiner A, Sahora A, Lachowicz J, Phillips B, Turner A, Klein MK, Post G, Diaz LA Jr, Riggins GJ, Papadopoulos N, Kinzler KW, Vogelstein B, Bettegowda C, Huso DL, Varterasian M, Saha S, Zhou S. Intratumoral injection of Clostridium novyi-NT spores induces antitumor responses. Sci Transl Med 2014;6:249ra111.
14. Toneri M, Miwa S, Zhang Y, Hu C, Yano S, Matsumoto Y, Bouvet M, Nakanishi H, Hoffman RM, Zhao M. Tumor-targeting Salmonella typhimurium A1-R inhibits human prostate cáncer experimental bone metástasis in mouse models. Oncotarget 2015;6:31335-43.
15. Laurence M, Yvonne P. Attenuated listeria monocytogenes: a powerful and versatile vector for the future of tumor immunotherapy. Front Cell Infect Microbiol 2014;4:51.
16. Gatenby R, Brown J. Mutations, evolution and the central role of a self-defined fitness function in the initiation and progression of cancer. Biochim Biophys Acta 2017;1867:162-6.
17. Zheng S, Widschwendter M, Teschendorff A. Epigenetic drift, epigenetic clocks and cancer risk. Epigenomics 2016;8:705-19.
18. Grasso Frisan T. Bacterial genotoxins: merging the DNA damage response into infection biology. Biomolecules 2015;5:1762-82.
20. Behrens A, van Deursen J, Rudolph K, Schumacher B. Impact of genomic damage and ageing on stem cell function. Nat Cell Biol 2014;16:201-7.
21. Charni M, Aloni-Grinstein R, Molchadsky A, Rotter V. p53 on the crossroad between regeneration and cancer. Cell Death Differ 2016;24:8-14.
22. Xu JH, Hu SL, Shen GD, Shen G. Tumor suppressor genesand their underlying interactions in paclitaxel resistance in cancer therapy. Cancer Cell Int 2016;16:13.
23. Shalapour S, Karin M. Immunity, inflammation, and cancer: an eternal fight between good and evil. J Clin Invest 2015;125:3347-55.
24. Garg A, Dudek A, Agostinis P. Cancer immunogenicity, danger signals, and DAMPs: what, when, and how? Biofactors 2013;39:355-67.
25. Pritchard A, Hastie M, Neller M, Gorman J, Schmidt C, Hayward N. Exploration of peptides bound to MHC class I molecules in melanoma. Pigment Cell Melanoma Res 2015;28:281-94.
26. Galon J, Angell H, Bedognetti D, Marincola F. The continuum of cancer immunosurveillance: prognostic, predictive, and mechanistic signatures. Immunity 2013;39:11-26.
27. Romee R, Leong J, Fehniger T. Utilizing cytokines to function-enable human NK cells for the immunotherapy of cancer. Scientifica (Cairo) 2014;2014:205796.
28. Gajewski T, Schreiber H, Fu Y. Innate and adaptive immune cells in the tumor microenvironment. Nat Immunol 2013;14:1014-22.
29. Payne K, Bear H, Manjili M. Adoptive cellular therapy of cancer: exploring innate and adaptive cellular crosstalk to improve anti-tumor efficacy. Future Oncol 2014;10:1779-94.
30. Gajewski TF, Schreiber H, Fu YX. Innate and adaptive immune cells in the tumor microenvironment. Nat Immunol 2013;14:1014-22.
31. Jandus C, Boligan K, Chijioke O, Liu H, Dahlhaus M, Démoulins T, Schneider C, Wehrli M, Hunger RE, Baerlocher GM, Simon HU, Romero P, Münz C, von Gunten S. Interactions between Siglec-7/9 receptors and ligands influence NK cell-dependent tumor immunosurveillance. J Clin Invest 2014;124:1810-20.
32. Morvan M, Lanier L. NK cells and cancer: you can teach innate cells new tricks. Nat Rev Cancer 2015;16:7-19.
33. Crouse J, Xu H, Lang P, Oxenius A. NK cells regulating T cell responses: mechanisms and outcome. Trends Immunol 2015;36:49-58.
34. D'Eliseo D, Manzi L, Velotti F. Capsaicin as an inducer of damage-associated molecular patterns (DAMPs) of immunogenic cell death (ICD) in human bladder cancer cells. Cell Stress Chaperones 2013;18:801-8.
35. Deauvieau F, Ollion V, Doffin A, Achard C, Fonteneau J, Verronese E, Durand I, Ghittoni R, Marvel J, Dezutter-Dambuyant C, Walzer T, Vie H, Perrot I, Goutagny N, Caux C, Valladeau-Guilemond J. Human natural killer cells promote cross-presentation of tumor cell-derived antigens by dendritic cells. Int J Cancer 2014;136:1085-94.
36. Ferlazzo G, Moretta L. Dendritic cell editing by natural killer cells. Crit Rev Oncog 2014;19:67-75.
37. Messina J, Fenstermacher D, Eschrich S, Qu X, Berglund A, Lloyd M, Schell MJ, Sondak VK, Weber JS, Mulé JJ. 12-Chemokine gene signature identifies lymph node-like structures in melanoma: potential for patient selection for immunotherapy? Sci Rep 2012;2:765.
38. Wilk E, Kalippke K, Buyny S, Schmidt R, Jacobs R. New aspects of NK cell subset identification and inference of NK cells' regulatory capacity by assessing functional and genomic profiles. Immunobiology 2008;213:271-83.
39. Della Chiesa M, Marcenaro E, Sivori S, Carlomagno S, Pesce S, Moretta A. Human NK cell response to pathogens. Semin Immunol 2014;26:152-60.
40. Slezak S, Worschech A, Wang E, Stroncek D, Marincola F. Analysis of vaccine-induced T cells in humans with cancer. Adv Exp Med Biol 2010;684:178-88.
41. Bhatia A, Kumar Y. Cancer stem cells and tumor immunoediting: putting two and two together. Expert Rev Clin Immunol 2016;12:605-7.
42. Calì B, Molon B, Viola A. Tuning cancer fate: the unremitting role of host immunity. Open Biol 2017;7:170006.
43. Mittal D, Gubin M, Schreiber R, Smyth M. New insights into cancer immunoediting and its three component phases-elimination, equilibrium and escape. Curr Opin Immunol 2014;27:16-25.
44. Wu X, Peng M, Huang B, Zhang H, Wang H, Huang B, Xue Z, Zhang L, Da Y, Yang D, Yao Z, Zhang R. Immune microenvironment profiles of tumor immune equilibrium and immune escape states of mouse sarcoma. Cancer Lett 2013;340:124-33.
45. Teng M, Galon J, Fridman W, Smyth M. From mice to humans: developments in cancer immunoediting. J Clin Invest 2015;125:3338-46.
46. Spranger S, Koblish H, Horton B, Scherle P, Newton R, Gajewski T. Mechanism of tumor rejection with doublets of CTLA-4, PD-1/PD-L1, or IDO blockade involves restored IL-2 production and proliferation of CD8+ T cells directly within the tumor microenvironment. J Immunother Cancer 2014;2:3.
47. Postow M, Callahan M, Wolchok J. Immune checkpoint blockade in cancer therapy. J Clin Oncol 2015;33:1974-82.
48. Vesely M, Schreiber R. Cancer immunoediting: antigens, mechanisms, and implications to cancer immunotherapy. Ann N Y Acad Sci 2013;1284:1-5.
49. Sakamoto C, Kohara H, Inoue H, Narusawa M, Ogawa Y, Hirose-Yotsuya L, Miyamoto S, Matsumura Y, Yamada K, Takahashi A, Tani K. Therapeutic vaccination based on side population cells transduced by the granulocyte-macrophage colony-stimulating factor gene elicits potent antitumor immunity. Cancer Gene Ther 2017;24:165-74.
51. Vedham V, Divi R, Starks V, Verma M. Multiple infections and cancer: implications in epidemiology. Technol Cancer Res Treat 2014;13:177-94.
52. Khan S, Imran A, Khan A, AbulKalam M, Alshamsan A. Systems biology approaches for the prediction of possible role of chlamydia pneumoniae proteins in the etiology of lung cancer. PLoS One 2016;11:e0148530.
53. Crowe S. Helicobacter infection, chronic inflammation, and the development of malignancy. Curr Opin Gastroenterol 2005;21:32-8.
54. Krishnan S, Eslick G. Streptococcus bovisinfection and colorectal neoplasia: a meta-analysis. Colorectal Dis 2014;16:672-80.
55. Hardbower D, de Sablet T, Chaturvedi R, Wilson K. Chronic inflammation and oxidative stress. Gut Microbes 2013;4:475-81.
56. Manda G, Isvoranu G, Comanescu M, Manea A, Debelec Butuner B, Korkmaz K. The redox biology network in cancer pathophysiology and therapeutics. Redox Biol 2015;5:347-57.
57. Kidane D, Chae WJ, Czochor J, Eckert KA, Glazer PM, Bothwell AL, Sweasy JB. Interplay between DNA repair and inflammation, and the link to cancer. Crit Rev Biochem Mol Biol 2014;49:116-39.
58. Hoesel B, Schmid J. The complexity of NF-κB signaling in inflammation and cancer. Mol Cancer 2013;12:86.
59. Elinav E, Nowarski R, Thaiss C, Hu B, Jin C, Flavell R. Inflammation-induced cancer: crosstalk between tumours, immune cells and microorganisms. Nat Rev Cancer 2013;13:759-71.
61. Van T, Polykratis A, Straub B, Kondylis V, Papadopoulou N, Pasparakis M. Kinase-independent functions of RIPK1 regulate hepatocyte survival and liver carcinogenesis. J Clin Invest 2017;127:2662-77.
62. Gambhir S, Vyas D, Hollis M, Aekka A, Vyas A. Nuclear factor kappa B role in inflammation associated gastrointestinal malignancies. World J Gastroenterol 2015;21:3174-83.
63. Pawlowski A, Jansson M, Sköld M, Rottenberg ME, Källenius G. Tuberculosis and HIV co-Infection. PLoS Pathog 2012;8:e1002464.
64. Scanu T, Spaapen R, Bakker J, Pratap C, Wu L, Hofland I, Broeks A, Shukla VK, Kumar M, Janssen H, Song JY, Neefjes-Borst EA, te Riele H, Holden DW, Nath G, Neefjes J. Salmonella manipulation of host signaling pathways provokes cellular transformation associated with gallbladder carcinoma. Cell Host Microbe 2015;17:763-74.
65. Zhang Y, Li S, Gan R, Zhou T, Xu D, Li H. Impacts of gut bacteria on human health and diseases. Int J Mol Sci 2015;16:7493-519.
66. Bernardes N, Chakrabarty A, Fialho A. Engineering of bacterial strains and their products for cancer therapy. Appl Microbiol Biotechnol 2013;97:5189-99.
67. Nauts H, Swift W, Coley B. The treatment of malignant tumors by bacterial toxins as developed by the late William B. Coley, MD, reviewed in the light of modern research. Cancer Res 1946;6:205-16.
68. Coley W. End results in Hodgkin's disease and lymphosarcoma treated by the mixed toxins of erysipelas and bacillus prodigiosus, alonis or combined with radiation. Ann Surg 1928;88:641.
69. Coley W. Treatment of inoperable malignant tumors with the toxines of erysipelas and the bacillus prodigiosus. Am J Med Sci 1894;108:50-66.
70. Coley W. The treatment of malignat tumors by repeated inoculations of erysipelas. Am J Med Sci 1893;105:487-510.
71. Coley W. The mixed toxins of erysipelas and bacillus prodigiosus in the treatment of sarcoma. JAMA 1900;XXXIV:906-8.
72. Nauts HC. The beneficial effects of bacterial infections on host resistance to cancer end results in 449 cases: a study and abstracts of reports in the world medical literature (1775-1980) and personal communications. Cancer Research Inst 1980;24:212-6.
73. Sell S. Cancer immunotherapy: breakthrough or "deja vu, all over again"? Tumor Biol 2017;39:1010428317707764.
74. Kienle G. Fever in cancer treatment: Coley's therapy and epidemiologic observations. Glob Adv Health Med 2012;1:92-100.
75. Rosenberg S. IL-2: the first effective immunotherapy for human cancer. J Immunol 2014;192:5451-8.
76. Le H, Lee N, Tsung K, Norton J. Pre-existing tumor-sensitized T cells are essential for eradication of established tumors by IL-12 and cyclophosphamide plus IL-12. J Immunol 2001;167:6765-72.
77. Kucerova P, Cervinkova M. Spontaneous regression of tumour and the role of microbial infection-possibilities for cancer treatment. Anticancer Drugs 2016;27:269-77.
78. Nallar S, Xu D, Kalvakolanu D. Bacteria and genetically modified bacteria as cancer therapeutics: current advances and challenges. Cytokine 2017;89:160-72.
79. Chorobik P, Czaplicki D, Ossysek K, Bereta J. Salmonella and cancer: from pathogens to therapeutics. Acta Biochim Pol 2013;60:285-97.
81. Brown K, Brown G, Lewis S, Beale R, Treacher D. Targeting cytokines as a treatment for patients with sepsis: a lost cause or a strategy still worthy of pursuit? Int Immunopharmacol 2016;36:291-9.
82. Dang LH, Bettegowda C, Huso DL, Kinzler KW, Vogelstein B. Combination bacteriolytic therapy for the treatment of experimental tumors. Proc Natl Acad Sci U S A 2001;98:15155-60.
83. Gujrati V, Kim S, Kim S, Min J, Choy H, Kim S, Jon S. Bioengineered bacterial outer membrane vesicles as cell-specific drug-delivery vehicles for cancer therapy. ACS Nano 2014;8:1525-37.
84. Piñero-Lambea C, Bodelón G, Fernández-Periáñez R, Cuesta A, Álvarez-Vallina L, Fernández L. Programming controlled adhesion of E. coli to target surfaces, cells, and tumors with synthetic adhesins. ACS Synth Biol 2015;4:463-73.
85. Lin I, Van T, Smooker P. Live-attenuated bacterial vectors: tools for vaccine and therapeutic agent delivery. Vaccines 2015;3:940-72.
86. Kasinskas R, Forbes N. Salmonella typhimurium lacking ribose chemoreceptors localize in tumor quiescence and induce apoptosis. Cancer Res 2007;67:3201-9.
87. Loeffler M, Le'Negrate G, Krajewska M, Reed J. Salmonella typhimurium engineered to produce CCL21 inhibit tumor growth. Cancer Immunol Immunother 2008;58:769-75.
88. Sahari A, Traore M, Scharf B, Behkam B. Directed transport of bacteria-based drug delivery vehicles: bacterial chemotaxis dominates particle shape. Biomed Microdevices 2014;16:717-25.
90. Alphandéry E. Applications of magnetosomes synthesized by magnetotactic bacteria in medicine. Front Bioeng Biotechnol 2014;2:5.
91. Pyne M, Bruder M, Moo-Young M, Chung D, Chou C. Technical guide for genetic advancement of underdeveloped and intractable Clostridium. Biotechnol Adv 2014;32:623-41.
92. Nallar S, Xu D, Kalvakolanu D. Bacteria and genetically modified bacteria as cancer therapeutics: current advances and challenges. Cytokine 2017;89:160-72.
93. Claesen J, Fischbach M. Synthetic microbes as drug delivery systems. ACS Synth Biol 2015;4:358-64.
94. Abil Z, Xiong X, Zhao H. Synthetic biology for therapeutic applications. Mol Pharm 2014;12:322-31.
95. Allen E, Miéville P, Warren CM, Saghafinia S, Li L, Peng MW, Hanahan D. Metabolic symbiosis enables adaptive resistance to anti-angiogenic therapy that is dependent on mTOR signaling. Cell Rep 2016;15:1144-60.
97. Phillips R. Targeting the hypoxic fraction of tumours using hypoxia-activated prodrugs. Cancer Chemother Pharmacol 2016;77:441-57.
98. Hunter F, Wouters B, Wilson W. Hypoxia-activated prodrugs: paths forward in the era of personalised medicine. Br J Cancer 2016;114:1071-7.
99. Rhim T, Lee D, Lee M. Hypoxia as a target for tissue specific gene therapy. J Control Release 2013;172:484-94.
100. Dhani N, Fyles A, Hedley D, Milosevic M. The clinical significance of hypoxia in human cancers. Semin Nucl Med 2015;45:110-21.
101. Heap J, Theys J, Ehsaan M, Kubiak A, Dubois L, Paesmans K, Van Mellaert L, Knox R, Kuehne SA, Lambin P, Minton NP. Spores of Clostridium engineered for clinical efficacy and safety cause regression and cure of tumors in vivo. Oncotarget 2014;5:1761-9.
102. Hammerich L, Brody JD. Immunomodulation within a single tumor site to induce systemic antitumor immunity: in situ vaccination for cancer. In: Rennert P, editor. Novel Immunotherapeutic Approaches to the Treatment of Cancer. Cham: Springer; 2016. pp. 129-62.
103. Park D, Park S, Cho S, Lee Y, Lee Y, Min J, Park BJ, Ko SY, Park JO, Park S. Motility analysis of bacteria-based microrobot (bacteriobot) using chemical gradient microchamber. Biotechnol Bioeng 2014;111:134-43.
104. Yu B, Shi L, Zhang BZ, Zhang KE, Peng X, Niu HB, Qu JL. Obligate anaerobic Salmonella typhimurium strain YB1 treatment on xenograft tumor in immunocompetent mouse model. Oncol Lett 2015;10:1069-74.
105. Thornlow D, Brackett E, Gigas J, Van Dessel N, Forbes N. Persistent enhancement of bacterial motility increases tumor penetration. Biotechnol Bioeng 2015;112:2397-405.
106. Pylaeva E, Lang S, Jablonska J. The essential role of type I interferons in differentiation and activation of tumor-associated neutrophils. Front Immunol 2016;7:629.
107. Hiroshima Y, Zhao M, Zhang Y, Zhang N, Maawy A, Murakami T, Mii S, Uehara F, Yamamoto M, Miwa S, Yano S, Momiyama M, Mori R, Matsuyama R, Chishima T, Tanaka K, Ichikawa Y, Bouvet M, Endo I, Hoffman RM. Tumor-targeting salmonella typhimurium A1-R arrests a chemo-resistant patient soft-tissue sarcoma in nude mice. PLoS One 2015;10:e0134324.
108. Matsumoto Y, Miwa S, Zhang Y, Hiroshima Y, Yano S, Uehara F, Yamamoto M, Toneri M, Bouvet M, Matsubara H, Hoffman RM, Zhao M. Efficacy of tumor-targetingsalmonella typhimuriumA1-R on nude mouse models of metastatic and disseminated human ovarian cancer. J Cell Biochem 2014;115:1996-2003.
109. Zoaby N, Shainsky-Roitman J, Badarneh S, Abumanhal H, Leshansky A, Yaron S, Schroeder A. Autonomous bacterial nanoswimmers target cancer. J Control Release 2017;257:68-75.
110. Toso J. Phase I study of the intravenous administration of attenuated salmonella typhimurium to patients with metastatic melanoma. J Clin Oncol 2002;20:142-52.
111. Zhang Y, Miwa S, Zhang N, Hoffman R, Zhao M. Tumor-targeting Salmonella typhimurium A1-R arrests growth of breast-cancer brain metastasis. Oncotarget 2014;6:2615-22.
112. Alimoradi H, Matikonda SS, Gamble AB, Giles GI, Greish K. Hypoxia responsive drug delivery systems in tumor therapy. Curr Pharm Des 2016;22:2808-20.
113. Loeffler M, Le'Negrate G, Krajewska M, Reed J. Attenuated Salmonella engineered to produce human cytokine LIGHT inhibit tumor growth. Proc Natl Acad Sci U S A 2007;104:12879-83.
114. Loeffler M, Le'Negrate G, Krajewska M, Reed J. IL-18-producing Salmonella inhibit tumor growth. Cancer Gene Ther 2008;15:787-94.
115. Loeffler M, Le'Negrate G, Krajewska M, Reed J. Inhibition of tumor growth using salmonella expressing fas ligand. J Natl Cancer Inst 2008;100:1113-6.
116. Ganai S, Arenas R, Forbes N. Tumour-targeted delivery of TRAIL using Salmonella typhimurium enhances breast cancer survival in mice. Br J Cancer 2009;101:1683-91.
117. Lee CH, Wu C, Shiau A. Endostatin gene therapy delivered bySalmonella choleraesuis in murine tumor models. J Gene Med 2004;6:1382-93.
118. Lee CH, Wu C, Shiau A. Salmonella choleraesuis as an anticancer agent in a syngeneic model of orthotopic hepatocellular carcinoma. Int J Cancer 2008;122:930-5.
119. Zhang L, Gao L, Zhao L, Guo B, Ji K, Tian Y, Wang J, Yu H, Hu J, Kalvakolanu DV, Kopecko DJ, Zhao X, Xu DQ. Intratumoral delivery and suppression of prostate tumor growth by attenuated salmonella enterica serovar typhimurium carrying plasmid-based small interfering RNAs. Cancer Res 2007;67:5859-64.
120. Niethammer AG, Xiang R, Becker J, Wodrich H, Pertl U, Karsten G, Eliceiri BP, Reisfeld RA. A DNA vaccine against VEGF receptor 2 prevents effective angiogenesis and inhibits tumor growth. Nat Med 2002;8:1369-75.
121. Fensterle J, Bergmann B, Yone C, Hotz C, Meyer S, Spreng S, Goebel W, Rapp UR, Gentschevl. Cancer immunotherapy based on recombinant Salmonella enterica serovar Typhimurium aroA strains secreting prostate-specific antigen and cholera toxin subunit B. Cancer Gene Ther 2007;15:85-93.
122. Lemmon MJ, van Zijl P, Fox ME, Mauchline ML, Giaccia AJ, Minton NP, Brown JM. Anaerobic bacteria as a gene delivery system that is controlled by the tumor microenvironment. Gene Ther 1997;4:791-6.
123. Fox ME, Lemmon MJ, Mauchline ML, Davis TO, Giaccia AJ, Minton NP, Brown JM. Anaerobic bacteria as a delivery system for cancer gene therapy: in vitro activation of 5-fluorocytosine by genetically engineered clostridia. Gene Ther 1996;3:173-8.
124. Zhang YL, Lü R, Chang ZS, Zhang WQ, Wang QB, Ding SY, Zhao W. Clostridium sporogenes delivers interleukin-12 to hypoxic tumours, producing antitumour activity without significant toxicity. Lett Appl Microbiol 2014;59:580-6.
125. Groot A, Mengesha A, Wall E, Diest P, Theys J, Vooijs M. Functional antibodies produced by oncolytic clostridia. Biochem Biophys Res Commun 2007;364:985-9.
126. Gunn GR, Zubair A, Peters C, Pan ZK, Wu TC, Paterson Y. Two listeria monocytogenes vaccine vectors that express different molecular forms of human papilloma virus-16 (HPV-16) E7 induce qualitatively different T cell immunity that correlates with their ability to induce regression of established tumors immortalized by HPV-16. J Immunol 2001;167:6471-9.
127. Shahabi V, Seavey M, Maciag P, Rivera S, Wallecha A. Development of a live and highly attenuated Listeria monocytogenes-based vaccine for the treatment of Her2/neu-overexpressing cancers in human. Cancer Gene Ther 2010;18:53-62.
128. Kim S, Castro F, Gonzalez D, Maciag P, Paterson Y, Gravekamp C. Mage-b vaccine delivered by recombinant Listeria monocytogenes is highly effective against breast cancer metastases. B J Cancer 2008;99:741-9.
129. Maciag PC, Seavey MM, Pan ZK, Ferrone S, Paterson Y. Cancer immunotherapy targeting the high molecular weight melanoma-associated antigen protein results in a broad antitumor response and reduction of pericytes in the tumor vasculature. Cancer Res 2008;68:8066-75.
130. Zu C, Wang J. Tumor-colonizing bacteria: a potential tumor targeting therapy. Crit Rev Microbiol 2013;40:225-35.
131. Quispe-Tintaya W, Chandra D, Jahangir A, Harris M, Casadevall A, Dadachova E, Gravekamp C. Nontoxic radioactive Listeriaat is a highly effective therapy against metastatic pancreatic cancer. Proc Natl Acad Sci U S A 2013;110:8668-73.
132. Hosseinidoust Z, Mostaghaci B, Yasa O, Park B, Singh A, Sitti M. Bioengineered and biohybrid bacteria-based systems for drug delivery. Adv Drug Deliv Rev 2016;106:27-44.
133. Fu W, Chu L, Han X, Liu X, Ren D. Synergistic antitumoral effects of human telomerase reverse transcriptase-mediated dual-apoptosis-related gene vector delivered by orally attenuated Salmonella enterica Serovar Typhimurium in murine tumor models. J Gene Med 2008;10:690-701.
134. Cicchelero L, Denies S, Devriendt B, de Rooster H, Sanders N. Can dendritic cells improve whole cancer cell vaccines based on immunogenically killed cancer cells? Onco Immunology 2015;4:e1048413.
135. Lin IY, Van TT, Smooker PM. Live-attenuated bacterial vectors: tools for vaccine and therapeutic agent delivery. Vaccines (Basel) 2015;3:940-72.
136. Inoue H, Tani K. Multimodal immunogenic cancer cell death as a consequence of anticancer cytotoxic treatments. Cell Death Differ 2013;21:39-49.
137. Tsuchiya K, Hara H, Fang R, Hernandez-Cuellar E, Sakai S, Daim S, Chen X, Dewamitta SR, Qu H, Mitsuyama M, Kawamura I. The adaptor ASC exacerbates lethal Listeria monocytogenes infection by mediating IL-18 production in an inflammasome-dependent and -independent manner. Eur J Immunol 2014;44:3696-707.
138. Srivastava S, Pelloso D, Feng H, Voiles L, Lewis D, Haskova Z, Whitacre M, Trulli S, Chen YJ, Toso J, Jonak ZL, Chang HC, Robertson MJ. Effects of interleukin-18 on natural killer cells: costimulation of activation through Fc receptors for immunoglobulin. Cancer Immunol Immunother 2013;62:1073-82.
139. Cao H, Xiang T, Zhang C, Yang H, Jiang L, Liu S, Huang X. MDA7 combined with targeted attenuated Salmonella vector SL7207/pBud-VP3 inhibited growth of gastric cancer cells. Biomed Pharmacother 2016;83:809-15.
140. Farooqi A, De Rosa G. TRAIL and microRNAs in the treatment of prostate cancer: therapeutic potential and role of nanotechnology. Appl Microbiol Biotechnol 2013;97:8849-57.
141. Zheng X, Ou Y, Shu M, Wang Y, Zhou Y, Su X, Zhu W, Yin W, Li S, Qiu P, Yan G, Zhang J, Hu J, Xu D. Cholera toxin, a typical protein kinase A activator, induces G1 phase growth arrest in human bladder transitional cell carcinoma cells via inhibiting the c-Raf/MEK/ERK signaling pathway. Mol Med Rep 2014;9:1773-9.
142. Zhao Q, Qu X, Liu K, Shi H, Yang G, Zhou B, Zhu L, Zhang W, Yan Z, Liu R, Qian S, Wang J. Microwave ablation combined with attenuated Salmonella typhimurium for treating hepatocellular carcinoma in a rat model. Oncotarget 2017;8:47655-64.
143. Morgan RA, Chinnasamy N, Abate-Daga D, Gros A, Robbins PF, Zheng Z, Dudley ME, Feldman SA, Yang JC, Sherry RM, Phan GQ, Hughes MS, Kammula US, Miller AD, Hessman CJ, Stewart AA, Restifo NP, Quezado MM, Alimchandani M, Rosenberg AZ, Nath A, Wang T, Bielekova B, Wuest SC, Akula N, McMahon FJ, Wilde S, Mosetter B, Schendel DJ, Laurencot CM, Rosenberg SA. .
144. Jiang SN, Phan TX, Nam TK, Nguyen VH, Kim HS, Bom HS, Choy HE, Hong Y, Min JJ. Inhibition of tumor growth and metastasis by a combination of Escherichia coli-mediated cytolytic therapy and radiotherapy. Mol Ther 2010;18:635-42.
145. Keates AC, Fruehauf J, Xiang S, Li CJ. TransKingdom RNA interference: a bacterial approach to challenges in RNAi therapy and delivery. Biotechnol Genet Eng Rev 2008;25:113-28.
146. Yang N, Li SH, Lü YZ, Chen LS, Ren DM. Attenuated Salmonella typhimurium carrying shRNA-expressing vectors elicit RNA interference in murine bladder tumors. Acta Pharmacol Sin 2011;32:368-74.
147. Tian Y, Guo B, Jia H, Ji K, Sun Y, Li Y, Zhao T, Gao L, Meng Y, Kalvakolanu DV, Kopecko DJ, Zhao X, Zhang L, Xu D. Targeted therapy via oral administration of attenuated Salmonella expression plasmid-vectored Stat3-shRNA cures orthotopically transplanted mouse HCC. Cancer Gene Ther 2012;19:393-401.
148. Yu YA, Weibel S, Szalay AA. Real-time imaging of tumors using replication-competent light-emitting microorganisms. Methods Mol Biol 2012;872:159-75.
149. Brader P, Stritzker J, Riedl CC, Zanzonico P, Cai S, Burnazi EM, Ghani ER, Hricak H, Szalay AA, Fong Y, Blasberg R. Escherichia coli Nissle 1917 facilitates tumor detection by positron emission tomography and optical imaging. Clin Cancer Res 2008;14:2295-302.
150. Viczián A, Kircher S. Luciferase and green fluorescent protein reporter genes as tools to determine protein abundance and intracellular dynamics. Methods Mol Biol 2010;655:293-312.
151. Voisey CR, Marincs F. Elimination of internal restriction enzyme sites from a bacterial luminescence (luxCDABE) operon. Biotechniques 1998;24:56-8.
152. Sörensen M, Lippuner C, Kaiser T, Misslitz A, Aebischer T, Bumann D. Rapidly maturing red fluorescent protein variants with strongly enhanced brightness in bacteria. FEBS Lett 2003;552:110-4.
153. Yu YA, Zhang Q, Szalay AA. Establishment and characterization of conditions required for tumor colonization by intravenously delivered bacteria. Biotechnol Bioeng 2008;100:567-78.
154. Araujo AC, Abreu F, Silva KT, Bazylinski DA, Lins U. Magnetotactic bacteria as potential sources of bioproducts. Mar Drugs 2015;13:389-430.
155. Nakamura C, Burgess JG, Sode K, Matsunaga T. An iron-regulated gene, magA, encoding an iron transport protein of magnetospirillum sp. strain AMB-1. J Biol Chem 1995;270:28392-6.
156. Mannucci S, Ghin L, Conti G, Tambalo S, Lascialfari A, Orlando T, Benati D, Bernardi P, Betterle N, Bassi R, Marzola P, Sbarbati A. Magnetic nanoparticles from magnetospirillum gryphiswaldense increase the efficacy of thermotherapy in a model of colon carcinoma. PLoS One 2014;9:e108959.
157. Want CZ, Kazmierczak RA, Eisenstark A. Straings, mechanism and perspective; Salmonella-based cancer therapy. Int J Microbiol 2016;2016:5678702.
158. Danino T, Prindle A, Kwong G, Skalak M, Li H, Allen K, Hasty J, Bhatia SN. Programmable probiotics for detection of cancer in urine. Sci Transl Med 2015;7:289ra84.
159. Mangtani P, Abubakar I, Ariti C, Beynon R, Pimpin L, Fine PE, Rodrigues LC, Smith PG, Lipman M, Whiting PF, Sterne JA. Protection by BCG vaccine against tuberculosis: a systematic review of randomized controlled trials. Clin Infect Dis 2013;58:470-80.
160. Kassouf W, Traboulsi SL, Kulkarni GS, Breau RH, Zlotta A, Fairey A, So A, Lacombe L, Rendon R, Aprikian AG, Siemens DR, Izawa JI, Black P. CUA guidelines on the management of non-muscle invasive bladder cancer. Can Urol Assoc J 2015;9:690.
161. Sylvester RJ, van der Meijden AP, Oosterlinck W, Witjes JA, Bouffioux C, Denis L, Newling DW, Kurth K. Predicting recurrence and progression in individual patients with stage Ta T1 bladder cancer using eortc risk tables: a combined analysis of 2596 patients from seven EORTC trials. Eur Urol 2006;49:466-77.
162. Sylvester RJ, Brausi MA, Kirkels WJ, Hoeltl W, Calais Da Silva F, Powell PH, Prescott S, Kirkali Z, van de Beek C, Gorlia T, de Reijke TM; EORTC Genito-Urinary Tract Cancer Group. Long-term efficacy results of eortc genito-urinary group randomized phase 3 study 30911 comparing intravesical instillations of epirubicin, bacillus calmette-guérin, and bacillus calmette-guérin plus isoniazid in patients with intermediate- and high-risk stage Ta T1 urothelial carcinoma of the bladder. Eur Urol 2010;57:766-73.
163. Babjuk M, Burger M, Zigeuner R, Shariat S, van Rhijn BW, Compérat E, Sylvester RJ, Kaasinen E, Böhle A, Palou Redorta J, Rouprêt M; European Association of Urology. EAU guidelines on non-muscle-invasive urothelial carcinoma of the bladder: update 2013. Eur Urol 2013;64:639-53.
164. Brausi M, Oddens J, Sylvester R, Bono A, van de Beek C, van Andel G, Gontero P, Turkeri L, Marreaud S, Collette S, Oosterlinck W. Side Effects of Bacillus Calmette-Guérin (BCG) in the treatment of intermediate- and high-risk Ta, T1 papillary carcinoma of the bladder: results of the EORTC genito-urinary cancers group randomised phase 3 study comparing one-third dose with full dose and 1 year with 3 years of maintenance BCG. Eur Urol 2014;65:69-76.
165. Redelman-Sidi G, Glickman M, Bochner B. The mechanism of action of BCG therapy for bladder cancer-a current perspective. Nat Rev Urol 2014;11:153-62.
166. Kawai K, Miyazaki J, Joraku A, Nishiyama H, Akaza H. Bacillus Calmette-Guerin (BCG) immunotherapy for bladder cancer: current understanding and perspectives on engineered BCG vaccine. Cancer Sci 2013;104:22-7.
167. Pandith AA, Khan NP, Azad NA, Khan MS, Wani MS. Association of chemokine and chemokine receptor gene polymorphis (MCP1 A-2518G and CCR2-V64I) with urinary bladder cancer: a study in kashmiri population. Am J Mol Cell Biol 2013;2:1-13.
168. Fuge O, Vasdev N, Allchorne P, Green JS. Immunotherapy for bladder cancer. Res Rep Urol 2015;7:65-79.
169. Bunimovich-Mendrazitsky S, Halachmi S, Kronik N. Improving Bacillus Calmette-Guérin (BCG) immunotherapy for bladder cancer by adding interleukin 2 (IL-2): a mathematical model. Math Med Biol 2016;33:159-88.
170. Iqbal NT, Hussain R. Non-specific immunity of BCG vaccine: a perspective of BCG immunotherapy. Trials Vaccinol 2014;3:143-9.
171. Kates M, Nirschl T, Sopko NA, Matsui H, Kochel CM, Reis LO, Netto GJ, Hoque MO, Hahn NM, McConkey DJ, Baras AS, Drake CG, Bivalacqua TJ. Intravesical BCG Induces CD4 + T-cell expansion in an immune competent model of bladder cancer. Cancer Immunol Res 2017;5:594-603.
172. Gandhi NM, Morales A, Lamm DL. Bacillus Calmette-Guérin immunotherapy for genitourinary cancer. BJU Int 2013;112:288-97.
173. Amarante-Mendes GP, Griffith TS. Therapeutic applications of TRAIL receptor agonists in cancer and beyond. Pharmacol Ther 2015;155:117-31.
174. Liu X, Dowell AC, Patel P, Viney RP, Foster MC, Porfiri E, James ND, Bryan RT. Cytokines as effectors and predictors of responses in the treatment of bladder cancer by bacillus Calmette-Guérin. Future Oncol 2014;10:1443-56.
175. Nemunaitis J, Cunningham C, Senzer N, Kuhn J, Cramm J, Litz C, Cavagnolo R, Cahill A, Clairmont C, Sznol M. Pilot trial of genetically modified, attenuated Salmonella expressing the E. coli cytosine deaminase gene in refractory cancer patients. Cancer Gene Ther 2003;10:737-44.
176. Schmitz-Winnenthal FH, Hohmann N, Niethammer A, Friedrich T, Lubenau H, Springer M, Breiner KM, Mikus G, Weitz J, Ulrich A, Buechler MW, Pianka F, Klaiber U, Diener M, Leowardi C, Schimmack S, Sisic L, Keller AV, Koc R, Springfeld C, Knebel P, Schmidt T, Ge Y, Bucur M, Stamova S, Podola L, Haefeli WE, Grenacher L, Beckhove P. Anti-angiogenic activity of VXM01, an oral T-cell vaccine against VEGF receptor 2, in patients with advanced pancreatic cancer: A randomized, placebo-controlled, phase 1 trial. Onco Immunol 2015;4:e1001217.
177. Le DT, Brockstedt D, Nir-Paz RG, Hampl J, Mathur S, Nemunaitis J, Sterman DH, Hassan R, Lutz E, Moyer B, Giedlin M, Louis JL, Sugar EA, Pons A, Cox AL, Levine J, Murphy AL, Illei P, Dubensky TW Jr, Eiden JE, Jaffee EM, Laheru DA. A live-attenuated listeria vaccine (ANZ-100) and a live-attenuated listeria vaccine expressing mesothelin (CRS-207) for advanced cancers: phase I studies of safety and immune induction. Clin Cancer Res 2011;18:858-68.
178. Le DT, Wang-Gillam A, Picozzi V, Greten TF, Crocenzi T, Springett G, Morse M, Zeh H, Cohen D, Fine RL, Onners B, Uram JN, Laheru DA, Lutz ER, Solt S, Murphy AL, Skoble J, Lemmens E, Grous J, Dubensky T Jr, Brockstedt DG, Jaffee EM. Safety and survival with GVAX pancreas prime and listeria monocytogenes-expressing mesothelin (CRS-207) boost vaccines for metastatic pancreatic cancer. J Clin Oncol 2015;33:1325-33.
179. Pollheimer M, Kornprat P, Lindtner R, Harbaum L, Schlemmer A, Rehak P, Langner C. Tumor necrosis is a new promising prognostic factor in colorectal cancer. Hum Pathol 2010;41:1749-57.
180. Ladstein RG, Bachmann IM, Straume O, Akslen LA. Tumor necrosis is a prognostic factor in thick cutaneous melanoma. Am J Surg Pathol 2012;36:1477-82.
181. Swinson D, Jones J, Richardson D, Cox G, Edwards J, O'Byrne K. Tumour necrosis is an independent prognostic marker in non-small cell lung cancer: correlation with biological variables. Lung Cancer 2002;37:235-40.
182. Staedtke V, Bai RY, Sun W, Huang J, Kibler KK, Tyler BM, Gallia GL, Kinzler K, Vogelstein B, Zhou S, Riggins GJ. Clostridium novyi-NT can cause regression of orthotopically implanted glioblastomas in rats. Oncotarget 2015;6:5536-46.
183. Krick EL, Sorenmo KU, Rankin SC, Cheong I, Kobrin B, Thornton K, Kinzler KW, Vogelstein B, Zhou S, Diaz LA Jr. Evaluation of Clostridium novyi -NT spores in dogs with naturally occurring tumors. Am J Vet Res 2012;73:112-8.
184. Kubiak A, Minton N. The potential of clostridial spores as therapeutic delivery vehicles in tumour therapy. Res Microbiol 2015;166:244-54.
185. Maciag P, Radulovic S, Rothman J. The first clinical use of a live-attenuated Listeria monocytogenes vaccine: a phase I safety study of Lm-LLO-E7 in patients with advanced carcinoma of the cervix. Vaccine 2009;27:3975-83.
186. Theys J, Pennington O, Dubois L, Anlezark G, Vaughan T, Mengesha A, Landuyt W, Anné J, Burke PJ, Dûrre P, Wouters BG, Minton NP, Lambin P. Repeated cycles of Clostridium-directed enzyme prodrug therapy result in sustained antitumour effects in vivo. Br J Cancer 2006;95:1212-9.
187. Fabricius EM, Schneeweiss U, Schau HP, Schmidt W, Benedix A. Quantitative investigations into the elimination of in vitro-obtained spores of the non-pathogenic Clostridium butyricum strain CNRZ 528, and their persistence in organs of different species following intravenous spore administration. Res Microbiol 1993;144:741-53.
188. Nuyts S, Van Mellaert L, Theys J, Landuyt W, Bosmans E, Anné J, Lambin P. Radio-responsive recA promoter significantly increases TNFα production in recombinant clostridia after 2 Gy irradiation. Gene Ther 2001;8:1197-1201.
189. Barbé S, Mellaert L, Theys J, Geukens N, Lammertyn E, Lambin P, Anné J. Secretory production of biologically active rat interleukin-2 by Clostridium acetobutylicum DSM792 as a tool for anti-tumor treatment. FEMS Microbiol Lett 2005;246:67-73.
190. Bermudes D, Low K, Ittensohn M, Le T, Platt J, Sodi S, Amoss M, Ash O, Carmichael E, Chakraborty A, Fischer J, Lin SL, Luo X, Miller SI, Zheng L, King I, Pawelek JM, Bermudes D. Lipid a mutant Salmonella with suppressed virulence and TNFalpha induction retain tumor-targeting in vivo. Nat Biotechnol 1999;17:37-41.
191. Clairmont C, Lee KC, Pike J, Ittensohn M, Low KB, Pawelek J, Bermudes D, Brecher SM, Margitich D, Turnier J, Li Z, Luo X, King I, Zheng LM. Biodistribution and genetic stability of the novel antitumor agent VNP20009, a genetically modified strain of Salmonella typhimurium. J Infect Dis 2000;181:1996-2002.
193. Aghi M, Hochberg F, Breakefield X. Prodrug activation enzymes in cancer gene therapy. J Gene Med 2000;2:148-64.
194. Nagakura C, Hayashi K, Zhao M, Yamauchi K, Yamamoto N, Tsuchiya H, Tomita K, Bouvet M, Hoffman RM. Efficacy of a genetically-modified Salmonella typhimurium in an orthotopic human pancreatic cancer in nude mice. Anticancer Res 2009;29:1873-8.
195. Hiroshima Y, Zhao M, Zhang Y, Maawy A, Hassanein M, Uehara F, Miwa S, Yano S, Momiyama M, Suetsugu A, Chishima T, Tanaka K, Bouvet M, Endo I, Hoffman RM. Comparison of efficacy of Salmonella typhimuriumA1-R and chemotherapy on stem-like and non-stem human pancreatic cancer cells. Cell Cycle 2013;12:2774-80.
196. Momiyama M, Zhao M, Kimura H, Tran B, Chishima T, Bouvet M, Endo I, Hoffman RM. Inhibition and eradication of human glioma with tumor-targetingSalmonella typhimuriumin an orthotopic nude-mouse model. Cell Cycle 2012;11:628-32.
197. Murakami T, Hiroshima Y, Zhao M, Zhang Y, Chishima T, Tanaka K, Bouvet M, Endo I, Hoffman RM. Therapeutic efficacy of tumor-targeting Salmonella typhimurium A1-R on human colorectal cancer liver metastasis in orthotopic nude-mouse models. Oncotarget 2015;6:31368-77.
198. Matsumoto Y, Miwa S, Zhang Y, Hiroshima Y, Yano S, Uehara F, Yamamoto M, Toneri M, Bouvet M, Matsubara H, Hoffman RM, Zhao M. Efficacy of tumor-targeting Salmonella typhimurium A1-R on nude mouse models of metastatic and disseminated human ovarian cancer. J Cell Biochem 2014;115:1996-2003.
199. Zhang Y, Zhang N, Zhao M, Hoffman RM. Comparison of the selective targeting efficacy of Salmonella typhimurium A1-R and VNP20009 on the Lewis lung carcinoma in nude mice. Oncotarget 2015;6:14625-31.
200. Yano S, Zhang Y, Miwa S, Tome Y, Hiroshima Y, Uehara F, Yamamoto M, Suetsugu A, Kishimoto H, Tazawa H, Zhao M, Bouvet M, Fujiwara T, Hoffman RM. Spatial-temporal FUCCI imaging of each cell in a tumor demonstrates locational dependence of cell cycle dynamics and chemoresponsiveness. Cell Cycle 2014;13:2110-9.
201. Yano S, Zhang Y, Zhao M, Hiroshima Y, Miwa S, Uehara F, Kishimoto H, Tazawa H, Bouvet M, Fujiwara T, Hoffman RM. Tumor-targeting Salmonella typhimurium A1-R decoys quiescent cancer cells to cycles as visualized by FUCCI imaging and become sensitive to chemotherapy. Cell Cycle 2014;13:2958-3963.
202. Jia H, Li Y, Zhao T, Li X, Hu J, Yin D, Guo B, Kopecko DJ, Zhao X, Zhang L, Xu DQ. Antitumor effects of Stat3-siRNA and endostatin combined therapies, delivered by attenuated Salmonella, on orthotopically implanted hepatocarcinoma. Cancer Immunol Immunother 2012;61:1977-87.
203. Park SY, Groisman EA. Signal-specific temporal response by the SalmonellaPhoP/PhoQ regulatory system. Mol Microbiol 2013;91:135-44.
204. Hohmann EL, Oletta CA, Killeen KP, Miller SI. PhoP/PhoQ-deleted Salmonella typhi (Ty800) is a safe and immunogenic single dose typhoid fever vaccine in volunteers. J Infect Dis 1996;173:1408-14.
205. Yu H, Pardoll D, Jove R. STATs in cancer inflammation and immunity: a leading role for STAT3. Nat Rev Cancer 2009;9:798-809.
206. Nam S, Buettner R, Turkson J, Kim D, Cheng JQ, Muehlbeyer S, Hippe F, Vatter S, Merz KH, Eisenbrand G, Jove R. Indirubin derivatives inhibit Stat3 signaling and induce apoptosis in human cancer cells. Proc Natl Acad Sci U S A 2005;102:5998-6003.
208. Chen Y, Liu X, Jin C, Zhou YC, Navab R, Jakobsen KR, Chen XQ, Li J, Li TT, Luo L, Wang XC. An orally administered DNA vaccine targeting vascular endothelial growth factor receptor-3 inhibits lung carcinoma growth. Tumour Biol 2015;37:2395-404.
209. Xu X, Hegazy WA, Guo L, Gao X, Courtney AN, Kurbanov S, Liu D, Tian G, Manuel ER, Diamond DJ, Hensel M, Metelitsa LS. Effective cancer vaccine platform based on attenuated Salmonella and a Type III secretion system. Cancer Res 2014;74:6260-70.
210. Weiskirch LM, Pan ZK, Paterson Y. The tumor recall response of antitumor immunity primed by a live, recombinant listeria monocytogenes vaccine comprises multiple effector mechanisms. Clin Immunol 2001;98:346-57.
211. Jahangir A, Chandra D, Quispe-Tintaya W, Singh M, Selvanesan BC, Gravekamp C. Immunotherapy with Listeria reduces metastatic breast cancer in young and old mice through different mechanisms. Onco Immunology 2017;6:e1342025.
212. Wallecha A, Singh R, Malinina I. Listeria monocytogenes (Lm)-LLO Immunotherapies reduce the immunosuppressive activity of myeloid-derived suppressor cells and regulatory T cells in the tumor microenvironment. J Immunother 2013;36:468-76.
213. Pan Z, Ikonomidis G, Pardoll D, Paterson Y. Regression of established tumors in mice mediated by the oral administration of a recombinant Listeria monocytogenes vaccine. Cancer Res 1995;55:4776-9.
214. Wallecha A, Carroll KD, Maciag PC, Rivera S, Shahabi V, Paterson Y. Multiple effector mechanisms induced by recombinant Listeria monocytogenes anticancer immunotherapeutics. Adv Appl Microbiol 2009;66:1-27.
215. Birmingham CL, Canadien V, Kaniuk NA, Steinberg BE, Higgins DE, Brumell JH. Listeriolysin O allows Listeria monocytogenes replication in macrophage vacuoles. Nature 2008;451:350-4.
216. Schnupf P, Zhou J, Varshavsky A, Portnoy DA. Listeriolysin O secreted by Listeria monocytogenes into the host cell cytosol is degraded by the N-end rule pathway. Infect Immun 2007;75:5135-47.
217. Eitel J, Suttorp N, Opitz B. Innate immune recognition and inflammasome activation in Listeria monocytogenes infection. Front Microbiol 2011;1:149.
218. Weiskirch L, Paterson Y. Listeria monocytogenes: a potent vaccine vector for neoplastic and infectious disease. Immunol Rev 1997;158:159-69.
219. Ikonomidis G, Paterson Y, Kos FJ, Portnoy DA. Delivery of a viral antigen to the class I processing and presentation pathway by Listeria monocytogenes. J Exp Med 1994;180:2209-18.
220. Mata M, Yao ZJ, Zubair A, Syres K, Paterson Y. Evaluation of a recombinant Listeria monocytogenes expressing an HIV protein that protects mice against viral challenge. Vaccine 2001;19:1435-45.
221. Singh R, Wallecha A. Cancer immunotherapy using recombinant Listeria monocytogenes: transition from bench to clinic. Hum Vaccin 2011;7:497-505.
222. Tilney LG, Portnoy DA. Actin filaments and the growth, movement, and spread of the intracellular bacterial parasite, Listeria monocytogenes. J Cell Biol 1989;109:1597-608.
223. Theisen E, Sauer JD. Listeria monocytogenesand the inflammasome: from cytosolic bacteriolysis to tumorimmunotherapy. Curr Top Microbiol Immunol 2016;397:133-60.
224. Cory L, Chu C. ADXS-HPV: a therapeutic Listeria vaccination targeting cervical cancers expressing the HPV E7 antigen. Hum Vaccin Immunother 2014;10:3190-5.
225. Shahabi V, Reyes-Reyes M, Wallecha A, Rivera S, Paterson Y, Maciag P. Development of a Listeria monocytogenes based vaccine against prostate cancer. Cancer Immunol Immunother 2008;57:1301-3.
226. Chen Y, Yang D, Li S, Gao Y, Jiang R, Deng L, Frankel FR, Sun B. Development of a Listeria monocytogenes-based vaccine against hepatocellular carcinoma. Oncogene 2011;31:2140-52.
227. Keenan B, Saenger Y, Kafrouni M, Leubner A, Lauer P, Maitra A, Rucki AA, Gunderson AJ, Coussens LM, Brockstedt DG, Dubensky TW Jr, Hassan R, Armstrong TD, Jaffee EM. A Listeria vaccine and depletion of T-regulatory cells activate immunity against early stage pancreatic intraepithelial neoplasms and prolong survival of mice. Gastroenterology 2014;146:1784-94.
228. Wood L, Paterson Y. Attenuated Listeria monocytogenes: a powerful and versatile vector for the future of tumor immunotherapy. Front Cell Infect Microbiol 2014;4:51.
229. Brockstedt D, Bahjat K, Giedlin M, Liu W, Leong M, Luckett W, Luckett W, Gao Y, Schnupf P, Kapadia D, Castro G, Lim JY, Sampson-Johannes A, Herskovits AA, Stassinopoulos A, Bouwer HG, Hearst JE, Portnoy DA, Cook DN, Dubensky TW Jr. Killed but metabolically active microbes: a new vaccine paradigm for eliciting effector T-cell responses and protective immunity. Nat Med 2005;11:853-60.
230. Li Z, Zhao X, Higgins DE, Frankel FR. Conditional lethality yields a new vaccine strain of Listeria monocytogenes for the induction of cell-mediated immunity. Infect Immun 2005;73:5065-73.
231. Lauer P, Chow M, Loessner M, Portnoy D, Calendar R. Construction, characterization, and use of two Listeria monocytogenes site-specific phage integration vectors. J Bacteriol 2002;184:4177-86.
232. Angelakopoulos H, Loock K, Sisul DM, Jensen ER, Miller JF, Hohmann EL. Safety and shedding of an attenuated strain of Listeria monocytogenes with a deletion of actA/plcB in adult volunteers: a dose escalation study of oral inoculation. Infect Immun 2002;70:3592-3601.
233. Verch T, Pan ZK, Paterson Y. Listeria monocytogenes-based antibiotic resistance gene-free antigen delivery system applicable to other bacterial vectors and DNA vaccines. Infect Immun 2004;72:6418-25.
234. Sacco JJ, Evans M, Harrington KJ, Man S, Powell N, Shaw RJ, Jones TM. Systemic listeriosis following vaccination with the attenuated Listeria monocytogenestherapeutic vaccine, ADXS11-001. Hum Vaccin Immunother 2015;12:1085-6.
235. Correction: mesothelin-specific chimeric antigen receptor mRNA-engineered T cells induce antitumor activity in solid malignancies. .
236. Hassan R, Laszik ZG, Lerner M, Raffeld M, Postier R, Brackett D. Mesothelin is overexpressed in pancreaticobiliary adenocarcinomas but not in normal pancreas and chronic pancreatitis. Am J Clin Pathol 2005;124:838-45.
237. He X, Wang L, Riedel H, Wang K, Yang Y, Dinu CZ, Rojanasakul Y. Mesothelin promotes epithelial-to-mesenchymal transition and tumorigenicity of human lung cancer and mesothelioma cells. Mol Cancer 2017;16:63.
238. Mkrtichyan M, Chong N, Abu Eid R, Wallecha A, Singh R, Rothman J, Khleif SN. Anti-PD-1 antibody significantly increases therapeutic efficacy of Listeria monocytogenes (Lm)-LLO immunotherapy. J Immunother Cancer 2013;1:15.
239. Han KJ, Lee NK, Park H, Paik HD. Anticancer and anti-inflammatory activity of probiotic lactococcus lactis NK34. J Microbiol Biotechnol 2015;25:1697-701.
240. Kim JY, Woo HJ, Kim YS, Kim KH, Lee HJ. Cell cycle dysregulation induced by cytoplasm of lactococcus lactis ssp. lactis in SNUC2A, a colon cancer cell line. Nutr Cancer 2003;46:197-201.
241. Kim SY, Kim JE, Lee KW, Lee HJ. Lactococcus lactis ssp. lactis inhibits the proliferation of SNU-1 human stomach cancer cells through induction of G0/G1 cell cycle arrest and apoptosis via p53 and p21 expression. Ann N Y Acad Sci 2009;1171:270-5.
242. Maletzki C, Linnebacher M, Kreikemeyer B, Emmrich J. Pancreatic cancer regression by intratumoral injection of live Streptococcus pyogenes in a syngeneic mouse model. Gut 2008;57:483-91.
243. Yin X, Yu B, Tang Z, He B, Ren J, Xiao X, Tang W. Bifidobacterium infantis-mediated HSV-TK/GCV suicide gene therapy induces both extrinsic and intrinsic apoptosis in a rat model of bladder cancer. Cancer Gene Ther 2012;20:77-81.
244. Bhatnagar PK, Awasthi A, Nomellini JF, Smit J, Suresh MR. Anti-tumor effects of the bacterium caulobacter crescentus in murine tumor models. Cancer Biol Ther 2006;5:485-91.
Cite This Article
Export citation file: BibTeX | RIS
OAE Style
Torres W, Lameda V, Olivar LC, Navarro C, Fuenmayor J, Pérez A, Mindiola A, Rojas M, Martínez MS, Velasco M, Rojas J, Bermudez V. Bacteria in cancer therapy: beyond immunostimulation. J Cancer Metastasis Treat 2018;4:4. http://dx.doi.org/10.20517/2394-4722.2017.49
AMA Style
Torres W, Lameda V, Olivar LC, Navarro C, Fuenmayor J, Pérez A, Mindiola A, Rojas M, Martínez MS, Velasco M, Rojas J, Bermudez V. Bacteria in cancer therapy: beyond immunostimulation. Journal of Cancer Metastasis and Treatment. 2018; 4: 4. http://dx.doi.org/10.20517/2394-4722.2017.49
Chicago/Turabian Style
Torres, Wheeler, Víctor Lameda, Luis Carlos Olivar, Carla Navarro, Jorge Fuenmayor, Adrián Pérez, Andres Mindiola, Milagros Rojas, María Sofía Martínez, Manuel Velasco, Joselyn Rojas, Valmore Bermudez. 2018. "Bacteria in cancer therapy: beyond immunostimulation" Journal of Cancer Metastasis and Treatment. 4: 4. http://dx.doi.org/10.20517/2394-4722.2017.49
ACS Style
Torres, W.; Lameda V.; Olivar LC.; Navarro C.; Fuenmayor J.; Pérez A.; Mindiola A.; Rojas M.; Martínez MS.; Velasco M.; Rojas J.; Bermudez V. Bacteria in cancer therapy: beyond immunostimulation. J. Cancer. Metastasis. Treat. 2018, 4, 4. http://dx.doi.org/10.20517/2394-4722.2017.49
About This Article
Copyright

Data & Comments
Data


 Cite This Article 60 clicks
Cite This Article 60 clicks

 Like This Article 39
likes
Like This Article 39
likes






Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.